Due to the enzyme's role in cholesterol biosynthesis, there is interest in lanosterol synthase inhibitors as potential cholesterol-reducing drugs, to complement existing statins.[12]
Structure
Lanosterol synthase is a two-domain monomeric protein[10] composed of two connected (α/α) barrel domains and three smaller β-structures. The enzyme active site is in the center of the protein, closed off by a constricted channel. Passage of the (S)-2,3-epoxysqualene substrate through the channel requires a change in protein conformation. In eukaryotes, a hydrophobic surface (6% of the total enzyme surface area) is the ER membrane-binding region.[12]
The enzyme contains five fingerprint regions containing Gln-Trp motifs, which are also present in the highly analogous bacterial enzyme squalene-hopene cyclase.[12] Residues of these fingerprint regions contain stacked sidechains which are thought to contribute to enzyme stability during the highly exergonic cyclization reactions catalyzed by the enzyme.[13]
Lanosterol synthase also catalyzes the cyclization of 2,3;22,23-diepoxysqualene to 24(S),25-epoxylanosterol,[14] which is later converted to 24(S),25-epoxycholesterol.[15] Since the enzyme affinity for this second substrate is greater than for the monoepoxy (S)-2,3-epoxysqualene, under partial inhibition conversion of 2,3;22,23-diepoxysqualene to 24(S),25-epoxylanosterol is favored over lanosterol synthesis.[16] This has relevance for disease prevention and treatment.
Mechanism
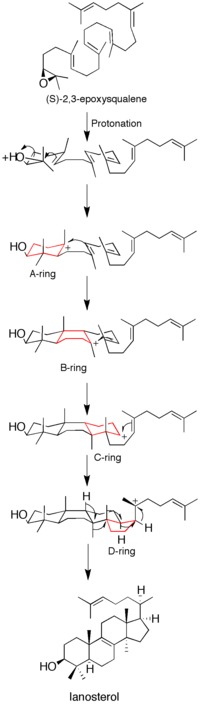
Figure 1: Lanosterol synthase mechanism. The discrete carbocation intermediates show the non-concerted nature of the mechanism.
Before the acquisition of the protein's X-ray crystal structure, site-directed mutagenesis was used to determine residues key to the enzyme's catalytic activity. It was determined that an aspartic acid residue (D455) and two histidine residues (H146 and H234) were essential to enzyme function. Corey et al. hypothesized that the aspartic acid acts by protonating the substrate's epoxide ring, thus increasing its susceptibility to intramolecular attack by the nearest double bond, with H146 possibly intensifying the proton donor ability of the aspartic acid through hydrogen bonding.[17] After acquisition of the X-ray crystal structure of the enzyme, the role of D455 as a proton donor to the substrate's epoxide was confirmed, though it was found that D455 is more likely stabilized by hydrogen bonding from two cysteine residues (C456 and C533) than from the earlier suggested histidine.[12]
Ring formation
Epoxide protonation activates the substrate, setting off a cascade of ring forming reactions. Four rings in total (A through D) are formed, producing the cholesterol backbone.[12] Though the idea of a concerted formation of all four rings had been entertained in the past, kinetic studies with (S)-2,3-oxidosqualene analogs showed that product formation is achieved through discrete carbocation intermediates (see Figure 1). Isolation of monocyclic and bicyclic products from lanosterol synthase mutants has further weakened the hypothesis of a concerted mechanism.[18][19] Evidence suggests that epoxide ring opening and A ring formation is concerted, though.[20]
Clinical significance
Enzyme inhibitors as cholesterol-lowering drugs
Interest has grown in lanosterol synthase inhibitors as drugs to lower blood cholesterol and treat atherosclerosis. The widely popular statin drugs currently used to lower LDL (low-density lipoprotein) cholesterol function by inhibiting HMG-CoA reductase activity.[6] Because this enzyme catalyzes the formation of precursors far upstream of (S)-2,3-epoxysqualene and cholesterol, statins may negatively influence amounts of intermediates required for other biosynthetic pathways (e.g. synthesis of isoprenoids, coenzyme Q). Thus, lanosterol synthase, which is more closely tied to cholesterol biosynthesis than HMG-CoA reductase, is an attractive drug target.[21]
Lanosterol synthase inhibitors are thought to lower LDL and VLDL cholesterol by a dual control mechanism. Studies in which lanosterol synthase is partially inhibited have shown both a direct decrease in lanosterol formation and a decrease in HMG-CoA reductase activity. The oxysterol 24(S),25-epoxylanosterol, which is preferentially formed over lanosterol during partial lanosterol synthase inhibition, is believed to be responsible for this inhibition of HMG-CoA reductase activity.[22]
Evolution
It is believed that oxidosqualene cyclases (OSCs, the class to which lanosterol cyclase belongs) evolved from bacterial squalene-hopene cyclase (SHC), which is involved with the formation of hopanoids. Phylogenetic trees constructed from the amino acid sequences of OSCs in diverse organisms suggest a single common ancestor, and that the synthesis pathway evolved only once.[23] The discovery of steranes including cholestane in 2.7-billion year-old shales from Pilbara Craton, Australia, suggests that eukaryotes with OSCs and complex steroid machinery were present early in earth's history.[24]
↑Baker CH, Matsuda SP, Liu DR, Corey EJ (Aug 1995). "Molecular cloning of the human gene encoding lanosterol synthase from a liver cDNA library". Biochemical and Biophysical Research Communications. 213 (1): 154–160. Bibcode:1995BBRC..213..154B. doi:10.1006/bbrc.1995.2110. PMID7639730.
↑Young M, Chen H, Lalioti MD, Antonarakis SE (May 1996). "The human lanosterol synthase gene maps to chromosome 21q22.3". Human Genetics. 97 (5): 620–624. doi:10.1007/BF02281872. PMID8655142. S2CID21051816.
123Ruf A, Müller F, D'Arcy B, Stihle M, Kusznir E, Handschin C, etal. (Mar 2004). "The monotopic membrane protein human oxidosqualene cyclase is active as monomer". Biochemical and Biophysical Research Communications. 315 (2): 247–254. Bibcode:2004BBRC..315..247R. doi:10.1016/j.bbrc.2004.01.052. PMID14766201.
↑Wendt KU, Poralla K, Schulz GE (Sep 1997). "Structure and function of a squalene cyclase". Science. 277 (5333). New York, N.Y.: 1811–1815. doi:10.1126/science.277.5333.1811. PMID9295270.
↑Corey EJ, Gross SK (Aug 1967). "Formation of sterols by the action of 2,3-oxidosqualene-sterol cyclase on the factitious substrates 2,3:22,23-dioxidosqualene and 2,3-oxido-22,23-dihydrosqualene". Journal of the American Chemical Society. 89 (17): 4561–4562. Bibcode:1967JAChS..89.4561C. doi:10.1021/ja00993a079. PMID6046552.
↑Boutaud O, Dolis D, Schuber F (Oct 1992). "Preferential cyclization of 2,3(S):22(S),23-dioxidosqualene by mammalian 2,3-oxidosqualene-lanosterol cyclase". Biochemical and Biophysical Research Communications. 188 (2): 898–904. Bibcode:1992BBRC..188..898B. doi:10.1016/0006-291X(92)91140-L. PMID1445330.
↑Corey EJ, Cheng CH, Baker CH, Matsuda SP, Li D, Song X (February 1997). "Studies on the Substrate Binding Segments and Catalytic Action of Lanosterol Synthase. Affinity Labeling with Carbocations Derived from Mechanism-Based Analogs of 2, 3-Oxidosqualene and Site-Directed Mutagenesis Probes". J. Am. Chem. Soc. 119 (6): 1289–1296. Bibcode:1997JAChS.119.1289C. doi:10.1021/ja963228o.
↑Wu TK, Wang TT, Chang CH, Liu YT, Shie WS (Nov 2008). "Importance of Saccharomyces cerevisiae oxidosqualene-lanosterol cyclase tyrosine 707 residue for chair-boat bicyclic ring formation and deprotonation reactions". Organic Letters. 10 (21): 4959–4962. doi:10.1021/ol802036c. PMID18842050.
↑Joubert BM, Hua L, Matsuda SP (Feb 2000). "Steric bulk at position 454 in Saccharomyces cerevisiae lanosterol synthase influences B-ring formation but not deprotonation". Organic Letters. 2 (3): 339–341. doi:10.1021/ol9912940. PMID10814317.
↑Corey EJ, Cheng CH, Baker CH, Matsuda SP, Li D, Song X (February 1997). "Methodology for the Preparation of Pure Recombinant S. cerevisiae Lanosterol Synthase Using a Baculovirus Expression System. Evidence That Oxirane Cleavage and A-Ring Formation Are Concerted in the Biosynthesis of Lanosterol from 2,3-Oxidosqualene". J. Am. Chem. Soc. 119 (6): 1277–1288. Bibcode:1997JAChS.119.1277C. doi:10.1021/ja963227w.
Young M, Chen H, Lalioti MD, Antonarakis SE (May 1996). "The human lanosterol synthase gene maps to chromosome 21q22.3". Human Genetics. 97 (5): 620–624. doi:10.1007/BF02281872. PMID8655142. S2CID21051816.
Peffley DM, Gayen AK, Morand OH (Aug 1998). "Down-regulation of 3-hydroxy-3-methylglutaryl coenzyme A reductase mRNA levels and synthesis in syrian hamster C100 cells by the oxidosqualene cyclase inhibitor [4'-(6-allyl-ethyl-amino-hexyloxy)-2'-fluoro-phenyl]-(4-bromophenyl)-me thanone (Ro 48-8071): comparison to simvastatin". Biochemical Pharmacology. 56 (4): 439–449. doi:10.1016/S0006-2952(98)00083-5. PMID9763219.
Pyrah IT, Kalinowski A, Jackson D, Davies W, Davis S, Aldridge A, etal. (2001). "Toxicologic lesions associated with two related inhibitors of oxidosqualene cyclase in the dog and mouse". Toxicologic Pathology. 29 (2): 174–179. doi:10.1080/019262301317052440. PMID11421484. S2CID38721319.
Ruf A, Müller F, D'Arcy B, Stihle M, Kusznir E, Handschin C, etal. (Mar 2004). "The monotopic membrane protein human oxidosqualene cyclase is active as monomer". Biochemical and Biophysical Research Communications. 315 (2): 247–254. Bibcode:2004BBRC..315..247R. doi:10.1016/j.bbrc.2004.01.052. PMID14766201.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.