This is a list of Antillian and Bermudan animals extinct in the Holocene that covers extinctions from the Holocene epoch, a geologic epoch that began about 11,650 years before present (about 9700 BCE) [a] and continues to the present day. [1] This list includes the Antilles archipelago and the Bermuda Islands, collectively known as the West Indies.
Contents
- Mammals (class Mammalia)
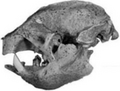
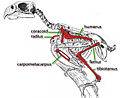
- Anteaters and sloths (order Pilosa)
- Rodents (order Rodentia)
- Primates (order Primates)
- True insectivores (order Eulipotyphla)
- Bats (order Chiroptera)
- Carnivorans (order Carnivora)
- Birds (class Aves)
- Nightjars (order Caprimulgiformes)
- Swifts, treeswifts, and hummingbirds (order Apodiformes)
- Pigeons and doves (order Columbiformes)
- Rails and cranes (order Gruiformes)
- Shorebirds (order Charadriiformes)
- Albatrosses and petrels (order Procellariiformes)
- Storks and allies (order Ciconiiformes)
- Pelicans, herons, and ibises (order Pelecaniformes)
- New World vultures (order Cathartiformes)
- Hawks and relatives (order Accipitriformes)
- Owls (order Strigiformes)
- Falcons (order Falconiformes)
- Woodpeckers and allies (order Piciformes)
- Parrots (order Psittaciformes)
- Perching birds (order Passeriformes)
- Reptiles (class Reptilia)
- Crocodilians (order Crocodilia)
- Squamates (order Squamata)
- Turtles and tortoises (order Testudines)
- Amphibians (class Amphibia)
- Frogs (order Anura)
- Insects (class Insecta)
- Butterflies (order Lepidoptera)
- Bark lice, book lice, and parasitic lice (order Psocodea)
- Arachnids (class Arachnida)
- Order Mesostigmata
- Order Sarcoptiformes
- Scorpions (order Scorpiones)
- Slugs and snails (class Gastropoda)
- Family Neocyclotidae
- Family Oleacinidae
- Family Pleurodontidae
- See also
- Notes
- References
The indigenous fauna of the West Indies collapsed in the Late Quaternary, with the rate of extinction for terrestrial mammals approaching 79-84%, one of the highest in the world. However, in stark contrast to the American continent, radiocarbon dating indicates that mammals survived the end of the Pleistocene with no apparent, or minimal losses despite localized sea level rise and climate change. [2] The same actually caused some bird extirpations and extinctions on the Bahamas, however. [3]
The indigenous peoples of the Caribbean arrived in the middle Holocene [4] and introduced guinea pigs, agoutis, and Native American dogs. While they accelerated the process, it was still not to the extent predicted by the "blitzkrieg" model of Pleistocene extinctions. For example, Caribbean sloths coexisted with humans for up to 400 years, even the largest species, which might indicate that they weren't commonly hunted. Some rodents, like the Puerto Rican hutia and Desmarest's hutia, and even flightless birds like the Antillean cave rail adapted well to human predation and were introduced to new islands by humans. [2] [5] European colonization, beginning with the arrival of Christopher Columbus in 1492, brought in Old World rats, mice, domestic animals, and large-scale deforestation that eradicated many of the native animals and those introduced by indigenous peoples alike. The most recent extinctions happened after the late 19th century, following the introduction of the small Indian mongoose for rat control. [2]
Many extinction dates are unknown due to a lack of relevant information.