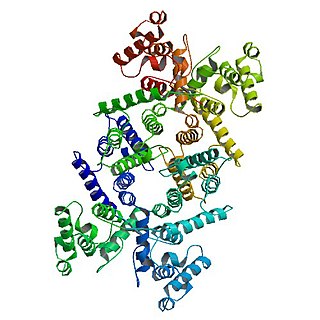
Dystrophin is a rod-shaped cytoplasmic protein, and a vital part of a protein complex that connects the cytoskeleton of a muscle fiber to the surrounding extracellular matrix through the cell membrane. This complex is variously known as the costamere or the dystrophin-associated protein complex (DAPC). Many muscle proteins, such as α-dystrobrevin, syncoilin, synemin, sarcoglycan, dystroglycan, and sarcospan, colocalize with dystrophin at the costamere. It has a molecular weight of 427 kDa

Duchenne muscular dystrophy (DMD) is a severe type of muscular dystrophy predominantly affecting boys. The onset of muscle weakness typically begins around age four, with rapid progression. Initially, muscle loss occurs in the thighs and pelvis, extending to the arms, which can lead to difficulties in standing up. By the age of 12, most individuals with Duchenne muscular dystrophy are unable to walk. Affected muscles may appear larger due to an increase in fat content, and scoliosis is common. Some individuals may experience intellectual disability, and females carrying a single copy of the mutated gene may show mild symptoms.

A Morpholino, also known as a Morpholino oligomer and as a phosphorodiamidate Morpholino oligomer (PMO), is a type of oligomer molecule used in molecular biology to modify gene expression. Its molecular structure contains DNA bases attached to a backbone of methylenemorpholine rings linked through phosphorodiamidate groups. Morpholinos block access of other molecules to small specific sequences of the base-pairing surfaces of ribonucleic acid (RNA). Morpholinos are used as research tools for reverse genetics by knocking down gene function.
The dystrophin-associated protein complex, also known as the dystrophin-associated glycoprotein complex is a multiprotein complex that includes dystrophin and the dystrophin-associated proteins. It is one of the two protein complexes that make up the costamere in striated muscle cells. The other complex is the integrin-vinculin-talin complex.
The miR-192 microRNA precursor, is a short non-coding RNA gene involved in gene regulation. miR-192 and miR-215 have now been predicted or experimentally confirmed in mouse and human.

The miR-129 microRNA precursor is a small non-coding RNA molecule that regulates gene expression. This microRNA was first experimentally characterised in mouse and homologues have since been discovered in several other species, such as humans, rats and zebrafish. The mature sequence is excised by the Dicer enzyme from the 5' arm of the hairpin. It was elucidated by Calin et al. that miR-129-1 is located in a fragile site region of the human genome near a specific site, FRA7H in chromosome 7q32, which is a site commonly deleted in many cancers. miR-129-2 is located in 11p11.2.
The miR-34 microRNA precursor family are non-coding RNA molecules that, in mammals, give rise to three major mature miRNAs. The miR-34 family members were discovered computationally and later verified experimentally. The precursor miRNA stem-loop is processed in the cytoplasm of the cell, with the predominant miR-34 mature sequence excised from the 5' arm of the hairpin.

MiR-155 is a microRNA that in humans is encoded by the MIR155 host gene or MIR155HG. MiR-155 plays a role in various physiological and pathological processes. Exogenous molecular control in vivo of miR-155 expression may inhibit malignant growth, viral infections, and enhance the progression of cardiovascular diseases.

In molecular biology, miR-137 is a short non-coding RNA molecule that functions to regulate the expression levels of other genes by various mechanisms. miR-137 is located on human chromosome 1p22 and has been implicated to act as a tumor suppressor in several cancer types including colorectal cancer, squamous cell carcinoma and melanoma via cell cycle control.

In molecular biology, mir-145 microRNA is a short RNA molecule that in humans is encoded by the MIR145 gene. MicroRNAs function to regulate the expression levels of other genes by several mechanisms.

In molecular biology miR-203 is a short non-coding RNA molecule. MicroRNAs function to regulate the expression levels of other genes by several mechanisms, such as translational repression and Argonaute-catalyzed messenger RNA cleavage. miR-203 has been identified as a skin-specific microRNA, and it forms an expression gradient that defines the boundary between proliferative epidermal basal progenitors and terminally differentiating suprabasal cells. It has also been found upregulated in psoriasis and differentially expressed in some types of cancer.

miR-296 is a family of microRNA precursors found in mammals, including humans. The ~22 nucleotide mature miRNA sequence is excised from the precursor hairpin by the enzyme Dicer. This sequence then associates with RISC which effects RNA interference.
In molecular biology, microRNAs function to regulate the expression levels of other genes by several mechanisms. The miR-449 microRNA family encompasses three homologous small RNA molecules (miR-449a/b/c). This miR-449 cluster is located in the second intron of the CDC20B gene which both are co-transcribed. This miR-449 family belongs to the miR-34/miR-449 superfamily of microRNAs that is composed of six homologous miRNAs, named miR-34a/b/c and miR-449a/b/c. They are grouped together in the same superfamily, as their seed region and their adjacent nucleotide sequences are largely conserved. The miR-449 miRNAs control the differentiation of multiciliated cells in vertebrates.
In molecular biology mir-497 microRNA is a short RNA molecule. MicroRNAs function to regulate the expression levels of other genes by several mechanisms.
In molecular biology mir-504 microRNA is a short RNA molecule. MicroRNAs function to regulate the expression levels of other genes by several mechanisms.
mir-612 microRNA is a short non-coding RNA molecule belonging both to the family of microRNAs and to that of small interfering RNAs (siRNAs). MicroRNAs function to regulate the expression levels of other genes by several mechanisms, whilst siRNAs are involved primarily with the RNA interference (RNAi) pathway. siRNAs have been linked through some members to the regulation of cancer cell growth, specifically in prostate adenocarcinoma.
In molecular biology mir-885 microRNA is a short RNA molecule. MicroRNAs function to regulate the expression levels of other genes by several mechanisms.

MiR-206 is a microRNA with a sequence conserved across most mammalian species, and in humans is a member of the myo-miR family of miRNAs, which includes miR-1, miR-133, and miR-208a/b. Mir-206 is well established for the regulation of cellular processes involving skeletal muscle development, as well as mitochondrial functioning. miR-206 is studied in C2C12 myoblast cells as this is a widely used model for the study of cellular differentiation of skeletal muscle. The biogenesis of miR-206 is unique in that the primary mature transcript is generated from the 3p arm of the precursor microRNA hairpin rather than the 5p arm. Currently, miR-206 has approximately twelve miRNA family members, and the cognate seed sequence of the miR-206 family is conserved across all twelve miRNA members.
miR-324-5p is a microRNA that functions in cell growth, apoptosis, cancer, epilepsy, neuronal differentiation, psychiatric conditions, cardiac disease pathology, and more. As a microRNA, it regulates gene expression through targeting mRNAs. Additionally, miR-324-5p is both an intracellular miRNA, meaning it is commonly found within the microenvironment of the cell, and one of several circulating miRNAs found throughout the body. Its presence throughout the body both within and external to cells may contribute to miR-324-5p's wide array of functions and role in numerous disease pathologies – especially cancer – in various organ systems.
