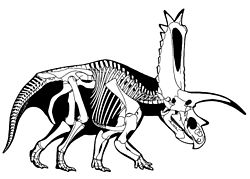
This timeline of ceratopsian research is a chronological listing of events in the history of paleontology focused on the ceratopsians, a group of herbivorous marginocephalian dinosaurs that evolved parrot-like beaks, bony frills, and, later, spectacular horns. The first scientifically documented ceratopsian fossils were described by Edward Drinker Cope starting in the 1870s; however, the remains were poorly preserved and their true nature was not recognized. Over the next several decades, Cope named several such genera and species. Cope's hated rival, Othniel Charles Marsh, also described ceratopsian remains. In 1887, Marsh mistook a Triceratops horn for one belonging to a new species of prehistoric Bison . [1] Marsh also named the eponymous genus Ceratops in 1888. [2] The next year, he named the most famous ceratopsian, Triceratops horridus . It was the discovery of Triceratops that illuminated the ceratopsian body plan, [1] and he formally named the Ceratopsia in 1890. [3]
Contents
- 19th century
- 1870s
- 1880s
- 1890s
- 20th century
- 1900s
- 1910s
- 1920s
- 1930s
- 1940s
- 1950s
- 1960s
- 1970s
- 1980s
- 1990s
- 21st century
- 2000s
- 2010s
- 2020s
- See also
- Footnotes
- References
The early 20th century was a fruitful time for ceratopsian research. In 1907, Hatcher and others published a monograph on ceratopsid anatomy that is still considered the single most significant publication on the topic to date. [1] Many new species were being described, including Centrosaurus , Styracosaurus , and Chasmosaurus . [4] Not long after, the Central Asiatic Expedition led by Roy Chapman Andrews of the American Museum of Natural History discovered the primitive ceratopsians Psittacosaurus and Protoceratops in Mongolia. Protoceratops gained notoriety as the supposed parent of the first fossil dinosaur eggs known to science. [5] One of these supposed Protoceratops nests preserved the skeleton of a new kind of theropod dinosaur, Oviraptor . It was thought to have been preserved after being killed in the act of raiding the Protoceratops nest. This narrative would last until the 1990s, when the "Protoceratops" eggs were determined to belong to Oviraptor itself. [6]
Throughout the rest of the century, paleontologists would be occupied with several controversies regarding ceratopsian paleobiology. One concerned the stance of the ceratopsid forelimbs. When Marsh first reconstructed the ceratopsid forelimb, he portrayed it in an erect posture. However, when later researchers like Sternberg and Osborn tried to mount the skeletons, they found that the forelimb bones apparently sprawled despite the hindlimbs standing straight up and down. Later researchers like Robert T. Bakker and Gregory S. Paul attempted to revive the erect reconstruction, but continuing research in the 1990s by researchers like John Ostrom, Peter Dodson, and James Farlow found an intermediate value to be better supported. [7]
The original use of the ceratopsids' horns and frills was another long-running controversy in ceratopsian paleontology. Early researchers like Richard Swann Lull thought that bony frills served as the attachment site for enlarged jaw muscles. This explanation was followed by researchers like Russell, Haas, and Ostrom. Sternberg thought the horns of ceratopsians helped defend against predators. In 1961, Davitashvili proposed that ceratopsids used their horns and frills to compete over mates. Farlow and Dodson arrived at the same conclusion in the 1970s, and were followed by Ralph Molnar. Ostrom, who had previously followed the jaw musculature interpretation, came to support this view in 1986. The idea gained further support in the 1990s from researchers like Forster and Sampson. [8]