Multituberculata is an extinct order of rodent-like mammals with a fossil record spanning over 130 million years. They first appeared in the Middle Jurassic, and reached a peak diversity during the Late Cretaceous and Paleocene. They eventually declined from the mid-Paleocene onwards, disappearing from the known fossil record in the late Eocene. They are the most diverse order of Mesozoic mammals with more than 200 species known, ranging from mouse-sized to beaver-sized. These species occupied a diversity of ecological niches, ranging from burrow-dwelling to squirrel-like arborealism to jerboa-like hoppers. Multituberculates are usually placed as crown mammals outside either of the two main groups of living mammals—Theria, including placentals and marsupials, and Monotremata—but usually as closer to Theria than to monotremes. They are considered to be closely related to Euharamiyida and Gondwanatheria as part of Allotheria.

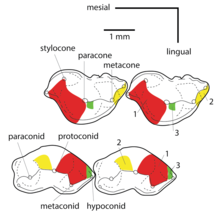
The molars or molar teeth are large, flat teeth at the back of the mouth. They are more developed in mammals. They are used primarily to grind food during chewing. The name molar derives from Latin, molaris dens, meaning "millstone tooth", from mola, millstone and dens, tooth. Molars show a great deal of diversity in size and shape across the mammal groups. The third molar of humans is sometimes vestigial.
Ferugliotherium is a genus of fossil mammals in the family Ferugliotheriidae from the Campanian and/or Maastrichtian period of Argentina. It contains a single species, Ferugliotherium windhauseni, which was first described in 1986. Although originally interpreted on the basis of a single brachydont (low-crowned) molar as a member of Multituberculata, an extinct group of small, rodent-like mammals, it was recognized as related to the hypsodont (high-crowned) Sudamericidae following the discovery of additional material in the early 1990s. After a jaw of the sudamericid Sudamerica was described in 1999, these animals were no longer considered to be multituberculates and a few fossils that were previously considered to be Ferugliotherium were assigned to unspecified multituberculates instead. Since 2005, a relationship between gondwanatheres and multituberculates has again received support. A closely related animal, Trapalcotherium, was described in 2009 on the basis of a single tooth.
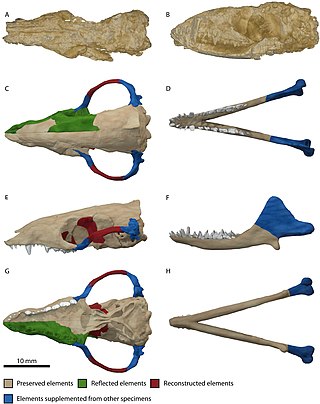
Morganucodon is an early mammaliaform genus that lived from the Late Triassic to the Middle Jurassic. It first appeared about 205 million years ago. Unlike many other early mammaliaforms, Morganucodon is well represented by abundant and well preserved material. Most of this comes from Glamorgan in Wales, but fossils have also been found in Yunnan Province in China and various parts of Europe and North America. Some closely related animals (Megazostrodon) are known from exquisite fossils from South Africa.
Ferugliotheriidae is one of three known families in the order Gondwanatheria, an enigmatic group of extinct mammals. Gondwanatheres have been classified as a group of uncertain affinities or as members of Multituberculata, a major extinct mammalian order. The best-known representative of Ferugliotheriidae is the genus Ferugliotherium from the Late Cretaceous epoch in Argentina. A second genus, Trapalcotherium, is known from a single tooth, a first lower molariform, from a different Late Cretaceous Argentinean locality. Another genus known from a single tooth, Argentodites, was first described as an unrelated multituberculate, but later identified as possibly related to Ferugliotherium. Finally, a single tooth from the Paleogene of Peru, LACM 149371, perhaps a last upper molariform, and a recent specimen from Mexico, may represent related animals.

Megazostrodon is an extinct genus of basal mammaliaforms belonging to the order Morganucodonta. It is approximately 200 million years old. Two species are known: M. rudnerae from the Early Jurassic of Lesotho and South Africa, and M. chenali from the Late Triassic of France.
Eozostrodon is an extinct morganucodont mammaliaform. It lived during the Rhaetian stage of the Late Triassic. Eozostrodon is known from disarticulated teeth from South West England and estimated to have been less than 10 cm (3.9 in) in head-body length, slightly smaller than the similar-proportioned Megazostrodon.

Symmetrodonta is a group of Mesozoic mammals and mammal-like synapsids characterized by the triangular aspect of the molars when viewed from above, and the absence of a well-developed talonid. The traditional group of 'symmetrodonts' ranges in age from the latest Triassic to the Late Cretaceous, but most research in the last 20-30 years has concluded that they are not a true taxonomic group, but include several unrelated branches of the mammal tree. Despite this, the name is still used informally by some researchers for convenience, usually restricted to the spalacotheriids and zhangheotheriids.
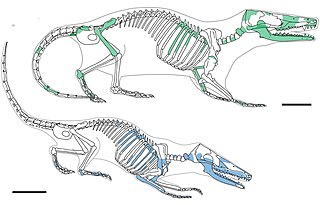
Docodonta is an order of extinct Mesozoic mammaliaforms. They were among the most common mammaliaforms of their time, persisting from the Middle Jurassic to the Early Cretaceous across the continent of Laurasia. They are distinguished from other early mammaliaforms by their relatively complex molar teeth. Docodont teeth have been described as "pseudotribosphenic": a cusp on the inner half of the upper molar grinds into a basin on the front half of the lower molar, like a mortar-and-pestle. This is a case of convergent evolution with the tribosphenic teeth of therian mammals. There is much uncertainty for how docodont teeth developed from their simpler ancestors. Their closest relatives may have been certain Triassic "symmetrodonts", namely Woutersia, Delsatia.

Mammaliaformes is a clade that contains the crown group mammals and their closest extinct relatives; the group radiated from earlier probainognathian cynodonts. It is defined as the clade originating from the most recent common ancestor of Morganucodonta and the crown group mammals; the latter is the clade originating with the most recent common ancestor of extant Monotremata, Marsupialia, and Placentalia. Besides Morganucodonta and the crown group mammals, Mammaliaformes includes Docodonta and Hadrocodium.
Sinoconodon is an extinct genus of mammaliamorphs that appears in the fossil record of the Lufeng Formation of China in the Sinemurian stage of the Early Jurassic period, about 193 million years ago. While sharing many plesiomorphic traits with other non-mammaliaform cynodonts, it possessed a special, secondarily evolved jaw joint between the dentary and the squamosal bones, which in more derived taxa would replace the primitive tetrapod one between the articular and quadrate bones. The presence of a dentary-squamosal joint is a trait historically used to define mammals.

The Australosphenida are a clade of mammals, containing mammals with tribosphenic molars, known from the Jurassic to Mid-Cretaceous of Gondwana. Although they have often been suggested to have acquired tribosphenic molars independently from those of Tribosphenida, this has been disputed. Fossils of australosphenidans have been found from the Jurassic of Madagascar and Argentina, and Cretaceous of Australia and Argentina. Monotremes have also been considered a part of this group in many studies, but this is also disputed.

Eutriconodonta is an order of early mammals. Eutriconodonts existed in Asia, Africa, Europe, North and South America during the Jurassic and the Cretaceous periods. The order was named by Kermack et al. in 1973 as a replacement name for the paraphyletic Triconodonta.

Morganucodonta is an extinct order of basal Mammaliaformes, a group including crown-group mammals (Mammalia) and their close relatives. Their remains have been found in Southern Africa, Western Europe, North America, India and China. The morganucodontans were probably insectivorous and nocturnal, though like eutriconodonts some species attained large sizes and were carnivorous. Nocturnality is believed to have evolved in the earliest mammals in the Triassic as a specialisation that allowed them to exploit a safer, night-time niche, while most larger predators were likely to have been active during the day.
Shuotherium is a fossil mammaliaform known from Middle-Late Jurassic of the Forest Marble Formation of England, and the Shaximiao Formation of Sichuan, China.
Yinotheria is a proposed basal subclass clade of crown mammals uniting the Shuotheriidae, an extinct group of mammals from the Jurassic of Eurasia, with Australosphenida, a group of mammals known from the Jurassic to Cretaceous of Gondwana, which possibly include living monotremes. Today, there are only five surviving species of monotremes which live in Australia and New Guinea, consisting of the platypus and four species of echidna. Fossils of yinotheres have been found in Britain, China, Russia, Madagascar and Argentina. Contrary to other known crown mammals, they retained postdentary bones as shown by the presence of a postdentary trough. The extant members (monotremes) developed the mammalian middle ear independently.
Doris Mary Kermack FLS was a British paleontologist and marine zoologist at Imperial College London. She completed her PhD thesis entitled 'The anatomy and physiology of the gut of Arenicola marina L.' at University College London in 1953. In 1988 she was awarded the Linnean Gold Medal for outstanding service to the society. She had two sons with her husband and fellow paleontologist Kenneth Kermack.
Kenneth Alexander Kermack was a British palaeontologist at University College London most notable for his work on early mammals with his wife, Doris Mary Kermack.

Triconodon is a genus of extinct mammal from the Early Cretaceous of England and France with two known species: T. mordax and T. averianovi. First described in 1859 by Richard Owen, it is the type genus for the order Triconodonta, a group of mammals characterised by their three-cusped (triconodont) molar teeth. Since then, this "simplistic" type of dentition has been understood to be either ancestral for mammals or else to have evolved multiple times, rendering "triconodonts" a paraphyletic or polyphyletic assemblage respectively, but several lineages of "triconodont" mammals do form a natural, monophyletic group, known as Eutriconodonta, of which Triconodon is indeed part of.
Vilevolodon is an extinct, monotypic genus of volant, arboreal euharamiyids from the Oxfordian age of the Late Jurassic of China. The type species is Vilevolodon diplomylos. The genus name Vilevolodon references its gliding capabilities, Vilevol, while don is a common suffix for mammalian taxon titles. The species name diplomylos refers to the dual mortar-and-pestle occlusion of upper and lower molars observed in the holotype; diplo, mylos.