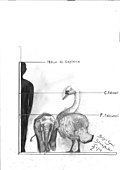
Size comparison of the giant gymnure/moonrat (a type of insectivore closely related to hedgehogs) Deinogalerix from the Late Miocene of Gargano, Italy, next to the related European hedgehog (Erinaceus europaeus)
Island gigantism, or insular gigantism, is a biological phenomenon in which the size of an animal species isolated on an island increases dramatically in comparison to its mainland relatives. Island gigantism is one aspect of the more general "island effect" or "Foster's rule", which posits that when mainland animals colonize islands, small species tend to evolve larger bodies, and large species tend to evolve smaller bodies (insular dwarfism). This is itself one aspect of the more general phenomenon of island syndrome which describes the differences in morphology, ecology, physiology and behaviour of insular (island) species compared to their continental counterparts. Following the arrival of humans and associated introduced predators (dogs, cats, rats, pigs), many giant as well as other island endemics have become extinct (e.g. the dodo and Rodrigues solitaire, giant flightless pigeons related to the Nicobar pigeon). A similar size increase, as well as increased woodiness, has been observed in some insular plants such as the Mapou tree (Cyphostemma mappia) in Mauritius which is also known as the "Mauritian baobab" although it is member of the grape family (Vitaceae).
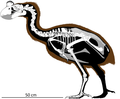
Diagram displaying the change in size of weta species in two ecosystems. The size and population of weta are affected by predation. Rats introduced on the mainland began to prey on weta, reducing their population; weta shrank in response. On an island isolated from predation, such as Little Barrier Island, weta have a dense population and have grown to a massive size. Insular species of giant weta are the only ones not facing extinction. As weta grow over time, bird predation declines.
Large mammalian carnivores are often absent on islands because of insufficient range or difficulties in over-water dispersal. In their absence, the ecological niches for large predators may be occupied by birds, reptiles or smaller carnivorans, which can then grow to larger-than-normal size. For example, on prehistoric Gargano Island in the Miocene-PlioceneMediterranean, on islands in the Caribbean like Cuba, and on Madagascar and New Zealand, some or all apex predators were birds like eagles, falcons and owls, including some of the largest known examples of these groups. However, birds and reptiles generally make less efficient large predators than advanced carnivorans.
Since small size usually makes it easier for herbivores to escape or hide from predators, the decreased predation pressure on islands can allow them to grow larger.[1][a] Small herbivores may also benefit from the absence of competition from missing types of large herbivores.
Benefits of large size that have been suggested for island tortoises include decreased vulnerability to scarcity of food and/or water, through ability to survive for longer intervals without them, or ability to travel longer distances to obtain them. Periods of such scarcity may be a greater threat on oceanic islands than on the mainland.[4]
Thus, island gigantism is usually an evolutionary trend resulting from the removal of constraints on the size of small animals related to predation and/or competition.[5] Such constraints can operate differently depending on the size of the animal, however; for example, while small herbivores may escape predation by hiding, large herbivores may deter predators by intimidation. As a result, the complementary phenomenon of island dwarfism can also result from the removal of constraints related to predation and/or competition on the size of large herbivores.[6] In contrast, insular dwarfism among predators more commonly results from the imposition of constraints associated with the limited prey resources available on islands.[6] As opposed to island dwarfism, island gigantism is found in most major vertebrate groups and in invertebrates.
Territorialism may favor the evolution of island gigantism. A study on Anaho Island in Nevada determined that reptile species that were territorial tended to be larger on the island compared to the mainland, particularly in the smaller species. In territorial species, larger size makes individuals better able to compete to defend their territory. This gives additional impetus to evolution toward larger size in an insular population.[7]
A further means of establishing island gigantism may be a founder effect operative when larger members of a mainland population are superior in their ability to colonize islands.[8]
Island size plays a role in determining the extent of gigantism. Smaller islands generally accelerate the rate of evolution of changes in organism size, and organisms there evolve greater extremes in size.[9]
The Komodo dragon of Flores and nearby islands, the largest extant lizard, and a similar (extinct) giant monitor lizard from Timor have been regarded as examples of giant insular carnivores. Since islands tend to offer limited food and territory, their mammalian carnivores (if present) are usually smaller than continental ones. These cases involve ectothermic carnivores on islands too small to support much mammalian competition. However, these lizards are not as large as their extinct Australian relative Megalania, and it has been proposed based on fossil evidence that the ancestors of these varanids first evolved their large size in Australia and then dispersed to Indonesia.[35] If this is true, rather than being insular giants they would be viewed as examples of phyletic gigantism. Supporting this interpretation is evidence for a lizard in Pliocene India, Varanus sivalensis, comparable in size to V. komodoensis.[35] Nevertheless, given that Australia is often described as the world's largest island and that the related Megalania, the largest terrestrial lizard known in the fossil record, was restricted to Australia, the perception of the largest Australasian/Indonesian lizards as insular giants may still have some validity.
Hatzegopteryx has features of island gigantism such as a more robust bodyplan and occupying niches taken by megafauna elsewhere (in this case, theropod dinosaurs).[41] However, similar sized giant pterosaurs occurred elsewhere, though nowhere near as robust.
In addition to size increase, island plants may also exhibit "insular woodiness".[49] The most notable examples are the megaherbs of New Zealand's subantarctic islands.[citation needed] Increased leaf and seed size was also reported in some island species regardless of growth form (herbaceous, bush, or tree).[50]
↑ The reduction in predation on islands often also leads to tamer behavior of island prey species, a trend that has been analyzed in lizards.[2][3]
↑ The earliest known New Zealand kiwi ancestor, a presumed recent arrival from Australia.[20]
↑ Based on the estimated total length of H. delcourti, ~23.6 in,[28] and the average length of a member of Diplodactylus, the most species-rich genus of Australian diplodactylid geckos, ~3.5 in.[29]
↑ Based on the average total length of the larger subspecies, R. l. leachianus, ~15.5 in,[30] and the average length of a member of Diplodactylus, the most species-rich genus of Australian diplodactylid geckos, ~3.5 in.[29]
↑ Based on the average mass of the larger subspecies, R. l. leachianus, ~240 g,[30] with the average weight of a member of Diplodactylus, the most species-rich genus of Australian diplodactylid geckos, ~4 g.[29]
↑ Based on the average female snout to vent length (SVL) of L. palmatus, ~96 mm,[43] with the average female SVL of L. rufus, ~80 mm.[44]
↑Keehn, J. E.; Nieto, N. C.; Tracy, C. R.; Gienger, C. M.; Feldman, C. R. (2013-08-27). "Evolution on a desert island: Body size divergence between the reptiles of Nevada's Anaho Island and the mainland around Pyramid Lake". Journal of Zoology. 291 (4): 269–278. doi:10.1111/jzo.12066.
↑Turvey, S. T. (2006). "A new genus and species of giant hutia (Tainotherium valei) from the Quaternary of Puerto Rico: an extinct arboreal quadruped?". Journal of Zoology. 270 (4): 585–594. doi:10.1111/j.1469-7998.2006.00170.x.
↑Torres-Roig, E.; Agustí, J.; Bover, P.; Alcover, J.A. (2017). "A new giant cricetine from the basal Pliocene of Mallorca (Balearic Islands, western Mediterranean): biostratigraphic nexus with continental mammal zones". Historical Biology. 31 (5): 559–573. doi:10.1080/08912963.2017.1377194. S2CID135302585.
↑Freudenthal, M. (1985). Cricetidae (Rodentia) from the Neogene of Gargano (Prov. of Foggia, Italy). Rijksmuseum van Geologie en Mineralogie.
↑Lerner, Heather R.L.; Mindell, David P. (2005). "Phylogeny of eagles, Old World vultures, and other Accipitridae based on nuclear and mitochondrial DNA". Molecular Phylogenetics and Evolution. 37 (2): 327–346. Bibcode:2005MolPE..37..327L. doi:10.1016/j.ympev.2005.04.010. PMID15925523.
↑Maca-Meyer, N.; Carranza, S.; Rando, J. C.; Arnold, E. N.; Cabrera, V. M. (2003-12-01). "Status and relationships of the extinct giant Canary Island lizard Gallotia goliath (Reptilia: Lacertidae), assessed using ancient mtDNA from its mummified remains". Biological Journal of the Linnean Society. 80 (4): 659–670. doi:10.1111/j.1095-8312.2003.00265.x.
↑Pérez-García, A.; Vlachos, E.; Arribas, A. (2017). "The last giant continental tortoise of Europe: A survivor in the Spanish Pleistocene site of Fonelas P-1". Palaeogeography, Palaeoclimatology, Palaeoecology. 470: 30–39. Bibcode:2017PPP...470...30P. doi:10.1016/j.palaeo.2017.01.011. hdl:11336/53105.
123Measey, G.J.; Vences, M.; Drewes, R.C.; Chiari, Y.; Melo, M.; Bourles, B. (2006). "Freshwater paths across the ocean: molecular phylogeny of the frog Ptychadena newtoni gives insights into amphibian colonization of oceanic islands". Journal of Biogeography. 34 (1): 7–20. doi:10.1111/j.1365-2699.2006.01589.x. S2CID17562846.
↑"Leptopelis palmatus". amphibiaweb.org. University of California, Berkeley. 2008. Retrieved 29 February 2020.
↑"Leptopelis rufus". amphibiaweb.org. University of California, Berkeley. 2008. Retrieved 29 February 2020.
↑Neither coconut crabs nor their relatives can swim beyond the larva stage, making the adults land animals in practice. Coconut crabs can weigh over 4 kg (9 pounds); the largest hermit crabs of the related genus Coenobita, C. brevimanus of coastal Africa and Asia, only reaches 230 grams (0.5 pounds).
GigantothermyLarge ectothermic animals more easily maintain constant body temperature
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.