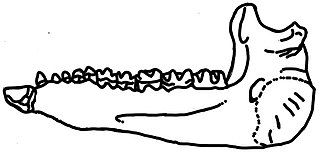
Forstercooperia is known from a vast amount of cranial material, although only some scant postcranial remains.[1] The average size of the genus is about equal with a large dog, even though later genera like Juxia and Paraceratherium reached sizes of a cow and even much larger.[2] Like primitive rhinocerotoids, Forstercooperia possesses blunt ends on the tips of its nasals, above the nasal incision. Unlike all modern rhinoceroses, the nasals of Forstercooperia, as well as many related genera, lack rugosities, which suggests that they lacked any form of horn. The nasal incision extends fairly far into the upper jaw, ending just posterior to the canine. Forstercooperia possesses a small post-insicor diastema, not as large as its descendants, and similar in size to that of Hyracodon.[3]
Taxonomy
Forstercooperia is considered to be a primitive rhinocerotoid, and as a result, many unrelated species were lumped into it. Species have also been oversplit based on small or insignificant features. In the most inclusive review of the genus yet, it was identified that many of the species are junior synonyms of previous names, not in the genus, or are potentially invalid in some other way. Many specimens, ranging in age from the Middle Eocene to Late Eocene, and in location from eastern Asia to Kazakhstan, and as far west as United States, have at some time been included in Forstercooperia. Material of many different genera as well have been at some time included in Forstercooperia, such as that of Juxia[1] and Uintaceras[4]
In 1923, a mostly complete skull of an early rhinoceros relative was unearthed. This skull came from the Late Eocene of the MongolianIrdin Manha Formation. The material, including the "front of skull with all premolars and some front teeth", was given the specimen numberAMNH 20116. It was first discovered in the formation by George Olsen, and in 1938 it was described as a new genus and species by Horace Elmer Wood II. Wood named the binomialCooperia totadentata to contain the skull. The generic name was to honor Clive Forster-Cooper, who had major contributions to the knowledge of indricotheres.[3] In 1939 Wood corrected the genus name to Forstercooperia, because he was informed that the name Cooperia was preoccupied, creating the new combination Forstercooperia totadentata.[5][1][6][7] The species is also the first named that has been included in Forstercooperia that is still valid.[4] It is also the senior synonym of F. shiwopuensis, a species named in 1974 by Chowet al..[1][8]
In 1963, material including a partial skull containing cheek teeth was unearthed in Late Eocene deposits of Mongolia. These remains were identified as from a true rhinoceros by Wood, who found them an important discovery with the scant amount of previous cranial material of early rhinocerotids available. On July 25, the same year, a paper was published by Wood concerning the taxonomy and osteology of these remains, in which he named them a new genus and species (or binomial) as well as re-ranking a previously named family as a subfamily containing the new taxon. The binomial created was Pappaceras confluens, classified as a close relative of Forstercooperia within Forstercooperiinae (before Forstercooperiidae, named in 1940 by Kretzoi). Wood noted that the generic name is derived from the Latin word πaππos, "grandfather", and the Greek words alpha, "without", and keras, "horn", translating as "Grandfather without horn". The species name is based on the confluent morphology of the teeth. The catalogue number for the skull is AMNH 26660, and it specifically preserved a "front half of the skull and a complete lower jaw, with most of the teeth and remaining alveoli, totaling a full placental series". Other remains included a portion of the mandible and a premolar. All of these specimens were from the lame locality, the Upper Gray Clays, of the Irdin Manha Formation in Inner Mongolia.[6] In the revision by Radnisky, it was found that this species was assignable to Forstercooperia, and the new combination F. confluens was erected.[9]
In the 1960s, newly uncovered material from the Irdin Manha Formation was identified as belonging to a new species of rhinocerotoid. Originally, they were found to be from F. confluens, as they were in the same location as that species holotype. They were later assigned to Forstercooperia sp., with no new name being given. The material included an almost complete skull, an almost complete lower jaw, an anterior portion of the skull, and an astragalus. These bones were first assigned a new species by Lucas et al., Forstercooperia minuta. They were found to be a unique species based on their size and the anatomy of their teeth. The species has been retained in the species complex of Forstercooperia throughout major revisions, by Lucas et al. in 1981,[1] Lucas and Sobus in 1989,[8] and Holbrook and Lucas in 1997. However, Holbrook and Lucas identified that the only North American material of F. minuta, F:AM 99662, had no features justifying its inclusion with the species, and reassigned it to their new binomial, Uintaceras radinskyi.[4]
An upper tooth row of an indricothere from the Eocene was first described in 1974. It was analysed by Chow, Chang and Ding, who published a new species for it, Forstercooperia shiwopuensis. The authors noted that it was from the same region and in the same size range as F. totadentata, which Lucas et al. (1981) found it to tentatively represent. The holotype of F. totadentata lacked an upper tooth row, and as it was presumably the right size to represent the missing teeth, Lucas et al. predicted it was of the same size and morphology as would have been predicted for the species.[1]
Forstercooperia has been represented by many different species in the different reviews of the genus.[1][4][8][9] In the first significant review, authored by Leonard Radinsky and published in 1967, found that many previous species were junior synonyms, and that only four species certainly in the genus were valid. Radinsky noted that of all published species, F. totadentata, F.? grandis, F. confluens, F. sharamurense, and F. borissiaki were the only valid ones, creating new combinations from Juxia sharamurense, Hyrachyus grandis, and Pappaceras borissiaki. He also synonymized the genera Pappaceras and Juxia with Forstercooperia.[9]
In a more recent review focused purely on the genus Forstercooperia, it was found that there was very little diversity in the species found valid by Radinsky. This paper, authored by Spencer G. Lucas and Robert Schoch and Earl Manning and published in 1981 reviewed all currently-named species of Forstercooperia, and named the new species F. minuta. Unlike Radinsky, their paper found Juxia to be separate, with F. borissiaki inside the genus, and F. grandis to be a definite species. F. confluens, named in 1963 by Wood, was found to be a synonym of F. grandis; F. sharamurunense, named in 1964 by Chow and Chiu, was found to be a synonym of F. borisiaki; F. jigniensis, named in 1973 by Sahni and Khare, was found to be an indeterminate species; F. shiwopuensis, named in 1974 by Chow, Chang and Ding, was found to be a synonym of F. totadentata; F. ergiliinensis, named in 1974 by Gabunia and Dashzeveg, was found to be a synonym of Juxia borissiaki; and F. crudus, named in 1977 by Gabunia, was found to be a nomen nudum.[1] In 1989, Lucas and Jay Sobus, it was noted that no new material of the genus had been identified, and that therefore, the conclusions were not changed.[8]
In the most recent review of the genus, focusing on the North American material, it was found that some earlier conclusions were no longer valid. This paper, published in 1997 by Luke Holbrook and Lucas, named a new genus, Uintaceras for all the North American material of Forstercooperia. Holbrook and Lucas named a new species, U. radinskyi, assigning the North American material of Fostercooperia to it. They found that the features uniting F. grandis with Forstercooperia were plesiomorphic, and that F. grandis was actually not a hyracodontid, instead the closest non-rhinocerotid relative of Rhinocerotidae. They concluded that F. grandis was a nomen dubium, its Asian material was assignable to F. confluens, Indricotheriinae was therefore only a Eurasian subfamily, and F. confluens was a valid species.[4] More recent mentions of Forstercooperia found no reason to contradict these conclusions.[10]
In 2016 new fostercooperiine material was reported from the early Eocene Arshanto Formation of Inner Mongolia, China. Wang and colleagues revalidated Pappaceras based on the new material and assigned it to a new species, P. meiomenus.[11]
The superfamilyRhinocerotoidea can be traced back to the early Eocene—about 50 million years ago—with early precursors such as Hyrachyus. Rhinocerotoidea contains three families; the Amynodontidae, the Rhinocerotidae ("true rhinoceroses"), and the Hyracodontidae. The diversity within the rhinoceros group was much larger in prehistoric times; sizes ranged from dog-sized to the size of Paraceratherium. There were long-legged, cursorial forms and squat, semi aquatic forms. Most species did not have horns. Rhinoceros fossils are identified as such mainly by characteristics of their teeth, which is the part of the animals most likely to be preserved. The upper molars of most rhinoceroses have a pi (π) shaped pattern on the crown, and each lower molar has paired L-shapes. Various skull features are also used for identification of fossil rhinoceroses.[2]
The subfamily Forstercooperiinae, to which Forstercooperia belongs, is considered part of Paraceratheriidae, a group containing the largest land mammals ever to walk the earth.[11] Paraceratheres are distinguished by their larger size and the derived structure of their snouts, incisors and canines. The earliest known indricothere is the dog-sized Pappaceras. The cow-sized Juxia is known from the middle Eocene; by the late Eocene the genus Urtinotherium of Asia had almost reached the size of the largest genus Paraceratherium.[2][8][4] The genus is distinct because of features of its nasal incision, dentition, tooth anatomy, and tooth proportions and size.[1] It retained the relatively primitive features of possessing three incisors, lower canines, and lower first premolars.[2]
Below is a phylogenetic analysis conducted by Lucas and Sobus in their 1989 revision of Indricotheriinae:[8]
In a 1999 study, Holbrook instead found the paraceratheres to be outside the hyracodontid group and wrote that the paraceratheres may not be a monophyletic grouping. He performed a phylogenetic analysis which placed Uintaceras, then amynodontids, then Paraceratherium, then Juxia and Forstercooperia, and finally hyracodontids as the successive outgroups of Rhinocerotidae within Rhinocerotoidea. The analysis however, did not include the genus Urtinotherium.[12] Later cladistic analysis confirmed some conclusions of Holbrook (1999), recovering hyracodontids as the basalmost rhinocerotoids and paraceratheres as closely related to Eggysodontidae and crown rhinos.[11]
Distribution and habitat
Location of finds of Forstercooperia
Remains of Forstercooperia have been found all across Asia. Most important remains are from the Middle to Late Eocene Irdin Manha Formation of Inner Mongolia (China).[1] In 1938, the holotype of F. totadentata was described, from the Irdin Manha Formation.[3] The only non-holotypic specimen of F. totadentata is that of F. shiwopuensis, which comes from the Lunan Formation of China.[1]
Related Research Articles
Camarasaurus was a genus of quadrupedal, herbivorous dinosaurs and is the most common North American sauropod fossil. Its fossil remains have been found in the Morrison Formation, dating to the Late Jurassic epoch, between 155 and 145 million years ago.
Andrewsarchus, meaning "Andrews' ruler", is an extinct genus of artiodactyl that lived during the Middle Eocene in what is now China. The genus was first described by Henry Fairfield Osborn in 1924 with the type species A. mongoliensis based on a largely complete cranium. A second species, A. crassum, was described in 1977 based on teeth. A mandible, formerly described as Paratriisodon, does probably belong to Andrewsarchus as well. The genus has been historically placed in the families Mesonychidae or Arctocyonidae, or was considered to be a close relative of whales. It is now regarded as the sole member of its own family, Andrewsarchidae, and may have been related to entelodonts. Fossils of Andrewsarchus have been recovered from the Middle Eocene Irdin Manha, Lushi, and Dongjun Formations of Inner Mongolia, each dated to the Irdinmanhan Asian land mammal age.
Paraceratherium is an extinct genus of hornless rhinocerotoids belonging to the family Paraceratheriidae. It is one of the largest terrestrial mammals that has ever existed and lived from the early to late Oligocene epoch. The first fossils were discovered in what is now Pakistan, and remains have been found across Eurasia between China and the Balkans. Paraceratherium means "near the hornless beast", in reference to Aceratherium, the genus in which the type species P. bugtiense was originally placed.
Paraceratheriidae is an extinct family of long-limbed, hornless rhinocerotoids, native to Asia and Eastern Europe that originated in the Eocene epoch and lived until the end of the Oligocene. They represent some of the largest terrestrial mammals to have ever lived.
Uintatherium is an extinct genus of herbivorous dinoceratan mammal that lived during the Eocene epoch. Two species are currently recognized: U. anceps from the United States during the Early to Middle Eocene and U. insperatus of Middle to Late Eocene China.
Amynodontidae is a family of extinct perissodactyls related to true rhinoceroses. They are commonly portrayed as semiaquatic hippo-like rhinos but this description only fits members of the Metamynodontini; other groups of amynodonts like the cadurcodontines had more typical ungulate proportions and convergently evolved a tapir-like proboscis.
The Hyracodontidae are an extinct family of rhinocerotoids endemic to North America, Europe, and Asia during the Eocene through early Oligocene, living from 48.6 to 26.3 million years ago (Mya), existing about 22.3 million years.
Juxia, ‘joo-she-a’, is an extinct genus of paraceratheriid, a group of herbivorous mammals that are related to the modern rhinoceros. The type species is Juxia sharamurenensis, named by Zhou Mingzhen and Qiu Zhanxiang in 1964. Juxia was around the size of a horse. It lived in Asia during the upper Eocene.
Brachyhyops is an extinct genus of entelodont artiodactyl mammal that lived during the Eocene Epoch of western North America and southeastern Asia. The first fossil remains of Brachyhyops are recorded from the late Eocene deposits of Beaver Divide in central Wyoming and discovered by paleontology crews from the Carnegie Museum of Natural History during the early 20th century. The type species, Brachyhyops wyomingensis, is based on a single skull and was named by E.H. Colbert in 1937, but was not officially described until 1938. During the latter half of the 20th century, additional specimens from North America have been recorded from Saskatchewan and as far south as Texas, indicating that Brachyhyops had a broad distribution and was well-dispersed throughout western North America.
Asiatosuchus is an extinct genus of crocodyloid crocodilians that lived in Eurasia during the Paleogene. Many Paleogene crocodilians from Europe and Asia have been attributed to Asiatosuchus since the genus was named in 1940. These species have a generalized crocodilian morphology typified by flat, triangular skulls. The feature that traditionally united these species under the genus Asiatosuchus is a broad connection or symphysis between the two halves of the lower jaw. Recent studies of the evolutionary relationships of early crocodilians along with closer examinations of the morphology of fossil specimens suggest that only the first named species of Asiatosuchus, A. grangeri from the Eocene of Mongolia, belongs in the genus. Most species are now regarded as nomina dubia or "dubious names", meaning that their type specimens lack the unique anatomical features necessary to justify their classification as distinct species. Other species such as "A." germanicus and "A." depressifrons are still considered valid species, but they do not form an evolutionary grouping with A. grangeri that would warrant them being placed together in the genus Asiatosuchus.
The Irdin Manha Formation is a geological formation from the Eocene located in Inner Mongolia, China, a few kilometres south of the Mongolian border.
Homogalax is an extinct genus of tapir-like odd-toed ungulate. It was described on the basis of several fossil finds from the northwest of the United States, whereby the majority of the remains come from the state of Wyoming. The finds date to the Lower Eocene between 56 and 48 million years ago. In general, Homogalax was very small, only reaching the weight of today's peccaries, with a maximum of 15 kg. Phylogenetic analysis suggests the genus to be a basal member of the clade that includes today's rhinoceros and tapirs. In contrast to these, Homogalax was adapted to fast locomotion.
Eggysodontidae is a family of perissodactyls, closely related to rhinoceroses. Fossils have been found in Oligocene deposits in Europe, the Caucasus, Central Asia, China, and Mongolia.
Urtinotherium is an extinct genus of paracerathere mammals. It was a large animal that was closely related to Paraceratherium, and found in rocks dating from the Late Eocene to Early Oligocene period. The remains were first discovered in the Urtyn Obo region in Inner Mongolia, which the name Urtinotherium is based upon. Other referred specimens are from northern China.
The Hsanda Gol Formation is a geological formation located in the Tsagan Nor Basin of central Mongolia that dates to the Rupelian age of the Oligocene epoch. It formed in continental desert-like conditions and is notable for its fossil mammals, first excavated by Roy Chapman Andrews and the American Museum of Natural History in the 1920s. Hsanda Gol is included in the Hsandagolian Asian Land Mammal Age, to which it gives its name.
Astorgosuchus is an extinct monospecific genus of crocodilian, closely related to true crocodiles, that lived in Pakistan during the late Oligocene period. This crocodile may have reached lengths of up to 7–8 m (23–26 ft) and is known to have preyed on many of the large mammals found in its environment. Bite marks of a large crocodile have been found on the bones of juvenile Paraceratherium, however if these were left by Astorgosuchus cannot be said with certainty. The genus contains a single species, Astorgosuchus bugtiensis, which was originally named as a species of Crocodylus in 1908 and was moved to its own genus in 2019.
Paracolodon is an extinct genus of tapiroid perissodactyl belonging to the family Helaletidae. Fossils have been found in Mongolia and the Inner Mongolia region of China.
Pappaceras is an extinct genus of rhinocerotoids from the Early Eocene of Asia belonging to Paraceratheriidae.
Dzungariotherium is a genus of paraceratheriid, an extinct group of large, hornless rhinocerotoids, which lived during the middle and late Oligocene of northwest China. The type species D. orgosense was described in 1973 based on fossils—mainly teeth—from Dzungaria in Xinjiang, northwest China.
Uintaceras is an extinct genus of medium-sized early rhinocerotoids that lived in North America during the Middle Eocene, with only the type species U. radinskyi, named in 1997, currently contained within the genus. Traditionally considered the oldest and most primitive species of the Rhinocerotidae, it may instead have been a close relative of the Asian Paraceratheriidae. The dubious species Forstercooperia (Hyrachyus) grandis is also possibly the same animal as Uintaceras, although the Asian material of F. grandis was assignable to Forstercooperia confluens.
1 2 3 4 5 6 7 8 9 10 11 12 Lucas, S.G.; Schoch, R.M.; Manning, E. (1981). "The Systematics of Forstercooperia, a Middle to Late Eocene Hyracodontid (Perissodactyla: Rhinocerotoidea) from Asia and Western North America". Journal of Paleontology. 55 (4): 826–841. JSTOR1304430.
1 2 3 4 Prothero, D.R. (2013). Rhinoceros Giants: The Palaeobiology of Indricotheres. Indiana University Press. pp.1–160. ISBN978-0-253-00819-0.
↑ Prothero, D.R. (2005). The Evolution of North American Rhinoceroses. Cambridge University Press. pp.1–218. ISBN0-521-83240-3.
1 2 3 Haibing Wang; Bin Bai; Jin Meng; Yuanqing Wang (2016). "Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids". Scientific Reports. 6: Article number 39607. doi:10.1038/srep39607.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.