
The hypothalamus is a small part of the brain that contains a number of nuclei with a variety of functions. One of the most important functions is to link the nervous system to the endocrine system via the pituitary gland. The hypothalamus is located below the thalamus and is part of the limbic system. It forms the ventral part of the diencephalon. All vertebrate brains contain a hypothalamus. In humans, it is the size of an almond.

The brainstem is the stalk-like part of the brain that interconnects the cerebrum and diencephalon with the spinal cord. In the human brain, the brainstem is composed of the midbrain, the pons, and the medulla oblongata. The midbrain is continuous with the thalamus of the diencephalon through the tentorial notch.

In neuroanatomy, the lateral geniculate nucleus is a structure in the thalamus and a key component of the mammalian visual pathway. It is a small, ovoid, ventral projection of the thalamus where the thalamus connects with the optic nerve. There are two LGNs, one on the left and another on the right side of the thalamus. In humans, both LGNs have six layers of neurons alternating with optic fibers.
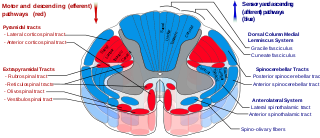
In anatomy, the extrapyramidal system is a part of the motor system network causing involuntary actions. The system is called extrapyramidal to distinguish it from the tracts of the motor cortex that reach their targets by traveling through the pyramids of the medulla. The pyramidal tracts may directly innervate motor neurons of the spinal cord or brainstem, whereas the extrapyramidal system centers on the modulation and regulation of anterior (ventral) horn cells.
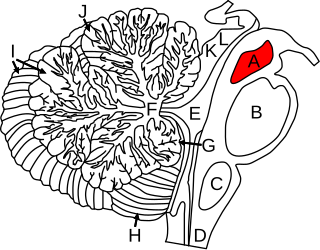
The midbrain or mesencephalon is the rostral-most portion of the brainstem connecting the diencephalon and cerebrum with the pons. It consists of the cerebral peduncles, tegmentum, and tectum.

The paraventricular nucleus is a nucleus in the hypothalamus. Anatomically, it is adjacent to the third ventricle and many of its neurons project to the posterior pituitary. These projecting neurons secrete oxytocin and a smaller amount of vasopressin, otherwise the nucleus also secretes corticotropin-releasing hormone (CRH) and thyrotropin-releasing hormone (TRH). CRH and TRH are secreted into the hypophyseal portal system and act on different targets neurons in the anterior pituitary. PVN is thought to mediate many diverse functions through these different hormones, including osmoregulation, appetite, and the response of the body to stress.
Magnocellular neurosecretory cells are large neuroendocrine cells within the supraoptic nucleus and paraventricular nucleus of the hypothalamus. They are also found in smaller numbers in accessory cell groups between these two nuclei, the largest one being the circular nucleus. There are two types of magnocellular neurosecretory cells, oxytocin-producing cells and vasopressin-producing cells, but a small number can produce both hormones. These cells are neuroendocrine neurons, are electrically excitable, and generate action potentials in response to afferent stimulation. Vasopressin is produced from the vasopressin-producing cells via the AVP gene, a molecular output of circadian pathways.

In neuroanatomy, the pretectal area, or pretectum, is a midbrain structure composed of seven nuclei and comprises part of the subcortical visual system. Through reciprocal bilateral projections from the retina, it is involved primarily in mediating behavioral responses to acute changes in ambient light such as the pupillary light reflex, the optokinetic reflex, and temporary changes to the circadian rhythm. In addition to the pretectum's role in the visual system, the anterior pretectal nucleus has been found to mediate somatosensory and nociceptive information.

The reticular formation is a set of interconnected nuclei that are located in the brainstem, hypothalamus, and other regions. It is not anatomically well defined, because it includes neurons located in different parts of the brain. The neurons of the reticular formation make up a complex set of networks in the core of the brainstem that extend from the upper part of the midbrain to the lower part of the medulla oblongata. The reticular formation includes ascending pathways to the cortex in the ascending reticular activating system (ARAS) and descending pathways to the spinal cord via the reticulospinal tracts.

The rubrospinal tract is a part of the nervous system. It is a part of the lateral indirect extra-pyramidal tract.
The red nucleus or nucleus ruber is a structure in the rostral midbrain involved in motor coordination. The red nucleus is pale pink, which is believed to be due to the presence of iron in at least two different forms: hemoglobin and ferritin. The structure is located in the tegmentum of the midbrain next to the substantia nigra and comprises caudal magnocellular and rostral parvocellular components. The red nucleus and substantia nigra are subcortical centers of the extrapyramidal motor system.

The globose nucleus is one of the deep cerebellar nuclei. It is located medial to the emboliform nucleus, and lateral to the fastigial nucleus. The globose nucleus and emboliform nucleus are known collectively as the interposed nuclei.
The zona incerta (ZI) is a horizontally elongated region of gray matter in the subthalamus below the thalamus. Its connections project extensively over the brain from the cerebral cortex down into the spinal cord.
The rubro-olivary tract is a tract which connects the inferior olive and the parvocellular red nucleus.
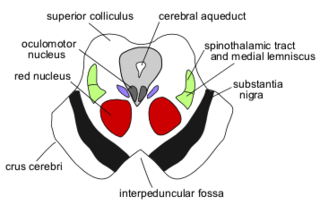
The central tegmental tract is a structure in the midbrain and pons.

The anatomy of the cerebellum can be viewed at three levels. At the level of gross anatomy, the cerebellum consists of a tightly folded and crumpled layer of cortex, with white matter underneath, several deep nuclei embedded in the white matter, and a fluid-filled ventricle in the middle. At the intermediate level, the cerebellum and its auxiliary structures can be broken down into several hundred or thousand independently functioning modules or compartments known as microzones. At the microscopic level, each module consists of the same small set of neuronal elements, laid out with a highly stereotyped geometry.
The parvocellular red nucleus (RNp) is located in the rostral midbrain and is involved in motor coordination. Together with the magnocellular red nucleus, it makes up the red nucleus.
Parvocellular neurosecretory cells are small neurons that produce hypothalamic releasing and inhibiting hormones. The cell bodies of these neurons are located in various nuclei of the hypothalamus or in closely related areas of the basal brain, mainly in the medial zone of the hypothalamus. All or most of the axons of the parvocellular neurosecretory cells project to the median eminence, at the base of the brain, where their nerve terminals release the hypothalamic hormones. These hormones are then immediately absorbed into the blood vessels of the hypothalamo-pituitary portal system, which carry them to the anterior pituitary gland, where they regulate the secretion of hormones into the systemic circulation.
The parabrachial nuclei, also known as the parabrachial complex, are a group of nuclei in the dorsolateral pons that surrounds the superior cerebellar peduncle as it enters the brainstem from the cerebellum. They are named from the Latin term for the superior cerebellar peduncle, the brachium conjunctivum. In the human brain, the expansion of the superior cerebellar peduncle expands the parabrachial nuclei, which form a thin strip of grey matter over most of the peduncle. The parabrachial nuclei are typically divided along the lines suggested by Baxter and Olszewski in humans, into a medial parabrachial nucleus and lateral parabrachial nucleus. These have in turn been subdivided into a dozen subnuclei: the superior, dorsal, ventral, internal, external and extreme lateral subnuclei; the lateral crescent and subparabrachial nucleus along the ventrolateral margin of the lateral parabrachial complex; and the medial and external medial subnuclei
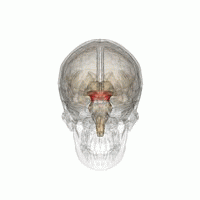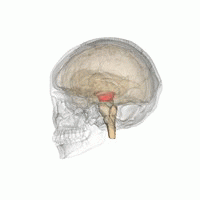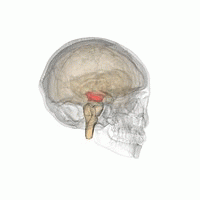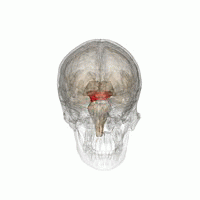
The mesencephalic locomotor region (MLR) is a functionally defined area of the midbrain that is associated with the initiation and control of locomotor movements in vertebrate species.
