The thalamus is a large mass of gray matter on the lateral walls of the third ventricle forming the dorsal part of the diencephalon. Nerve fibers project out of the thalamus to the cerebral cortex in all directions, known as the thalamocortical radiations, allowing hub-like exchanges of information. It has several functions, such as the relaying of sensory and motor signals to the cerebral cortex and the regulation of consciousness, sleep, and alertness.
Articles related to anatomy include:

The brainstem is the stalk-like part of the brain that interconnects the cerebrum and diencephalon with the spinal cord. In the human brain, the brainstem is composed of the midbrain, the pons, and the medulla oblongata. The midbrain is continuous with the thalamus of the diencephalon through the tentorial notch.

The visual system is the physiological basis of visual perception. The system detects, transduces and interprets information concerning light within the visible range to construct an image and build a mental model of the surrounding environment. The visual system is associated with the eye and functionally divided into the optical system and the neural system.

In neuroanatomy, the lateral geniculate nucleus is a structure in the thalamus and a key component of the mammalian visual pathway. It is a small, ovoid, ventral projection of the thalamus where the thalamus connects with the optic nerve. There are two LGNs, one on the left and another on the right side of the thalamus. In humans, both LGNs have six layers of neurons alternating with optic fibers.
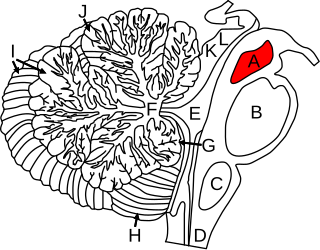
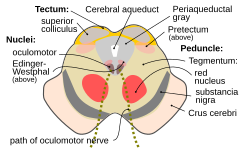
The midbrain or mesencephalon is the rostral-most portion of the brainstem connecting the diencephalon and cerebrum with the pons. It consists of the cerebral peduncles, tegmentum, and tectum.

The pupillary light reflex (PLR) or photopupillary reflex is a reflex that controls the diameter of the pupil, in response to the intensity (luminance) of light that falls on the retinal ganglion cells of the retina in the back of the eye, thereby assisting in adaptation of vision to various levels of lightness/darkness. A greater intensity of light causes the pupil to constrict, whereas a lower intensity of light causes the pupil to dilate. Thus, the pupillary light reflex regulates the intensity of light entering the eye. Light shone into one eye will cause both pupils to constrict.

The fibers of the oculomotor nerve arise from a nucleus in the midbrain, which lies in the gray substance of the floor of the cerebral aqueduct and extends in front of the aqueduct for a short distance into the floor of the third ventricle. From this nucleus the fibers pass forward through the tegmentum, the red nucleus, and the medial part of the substantia nigra, forming a series of curves with a lateral convexity, and emerge from the oculomotor sulcus on the medial side of the cerebral peduncle.

In neuroanatomy, the superior colliculus is a structure lying on the roof of the mammalian midbrain. In non-mammalian vertebrates, the homologous structure is known as the optic tectum or optic lobe. The adjective form tectal is commonly used for both structures.

The inferior colliculus (IC) is the principal midbrain nucleus of the auditory pathway and receives input from several peripheral brainstem nuclei in the auditory pathway, as well as inputs from the auditory cortex. The inferior colliculus has three subdivisions: the central nucleus, a dorsal cortex by which it is surrounded, and an external cortex which is located laterally. Its bimodal neurons are implicated in auditory-somatosensory interaction, receiving projections from somatosensory nuclei. This multisensory integration may underlie a filtering of self-effected sounds from vocalization, chewing, or respiration activities.

The reticular formation is a set of interconnected nuclei that are located in the brainstem, hypothalamus, and other regions. It is not anatomically well defined, because it includes neurons located in different parts of the brain. The neurons of the reticular formation make up a complex set of networks in the core of the brainstem that extend from the upper part of the midbrain to the lower part of the medulla oblongata. The reticular formation includes ascending pathways to the cortex in the ascending reticular activating system (ARAS) and descending pathways to the spinal cord via the reticulospinal tracts.
The Edinger–Westphal nucleus is one of two nuclei of the oculomotor nerve and is located in the midbrain. It receives afferents from both pretectal nuclei. It contains parasympathetic pre-ganglionic neuron cell bodies that synapse in the ciliary ganglion. It contributes the autonomic, parasympathetic component to the oculomotor nerve, ultimately providing innervation to the iris sphincter muscle and ciliary muscle to mediate the pupillary light reflex and accommodation, respectively.
Intrinsically photosensitive retinal ganglion cells (ipRGCs), also called photosensitive retinal ganglion cells (pRGC), or melanopsin-containing retinal ganglion cells (mRGCs), are a type of neuron in the retina of the mammalian eye. The presence of an additional photoreceptor was first suspected in 1927 when mice lacking rods and cones still responded to changing light levels through pupil constriction; this suggested that rods and cones are not the only light-sensitive tissue. However, it was unclear whether this light sensitivity arose from an additional retinal photoreceptor or elsewhere in the body. Recent research has shown that these retinal ganglion cells, unlike other retinal ganglion cells, are intrinsically photosensitive due to the presence of melanopsin, a light-sensitive protein. Therefore, they constitute a third class of photoreceptors, in addition to rod and cone cells.
The zona incerta (ZI) is a horizontally elongated region of gray matter in the subthalamus below the thalamus. Its connections project extensively over the brain from the cerebral cortex down into the spinal cord.
The optokinetic reflex (OKR), also referred to as the optokinetic response, or optokinetic nystagmus (OKN), is a compensatory reflex that supports visual image stabilization. The purpose of OKR is to prevent image blur on the retina that would otherwise occur when an animal moves its head or navigates through its environment. This is achieved by the reflexive movement of the eyes in the same direction as image motion, so as to minimize the relative motion of the visual scene on the eye. OKR is best evoked by slow, rotational motion, and operates in coordination with several complementary reflexes that also support image stabilization, including the vestibulo-ocular reflex (VOR).

The medial dorsal nucleus is a large nucleus in the thalamus.
The isothalamus is a division used by some researchers in describing the thalamus.
The parabrachial nuclei, also known as the parabrachial complex, are a group of nuclei in the dorsolateral pons that surrounds the superior cerebellar peduncle as it enters the brainstem from the cerebellum. They are named from the Latin term for the superior cerebellar peduncle, the brachium conjunctivum. In the human brain, the expansion of the superior cerebellar peduncle expands the parabrachial nuclei, which form a thin strip of grey matter over most of the peduncle. The parabrachial nuclei are typically divided along the lines suggested by Baxter and Olszewski in humans, into a medial parabrachial nucleus and lateral parabrachial nucleus. These have in turn been subdivided into a dozen subnuclei: the superior, dorsal, ventral, internal, external and extreme lateral subnuclei; the lateral crescent and subparabrachial nucleus along the ventrolateral margin of the lateral parabrachial complex; and the medial and external medial subnuclei
Lateral pulvinar nucleus is one of four traditionally anatomically distinguished nuclei of the pulvinar of the thalamus. The other three nuclei of the pulvinar are called anterior, inferior and medial pulvinar nuclei.
Medial pulvinar nucleus is one of four traditionally anatomically distinguished nuclei of the pulvinar of the thalamus. The other three nuclei of the pulvinar are called lateral, inferior and anterior pulvinar nuclei.