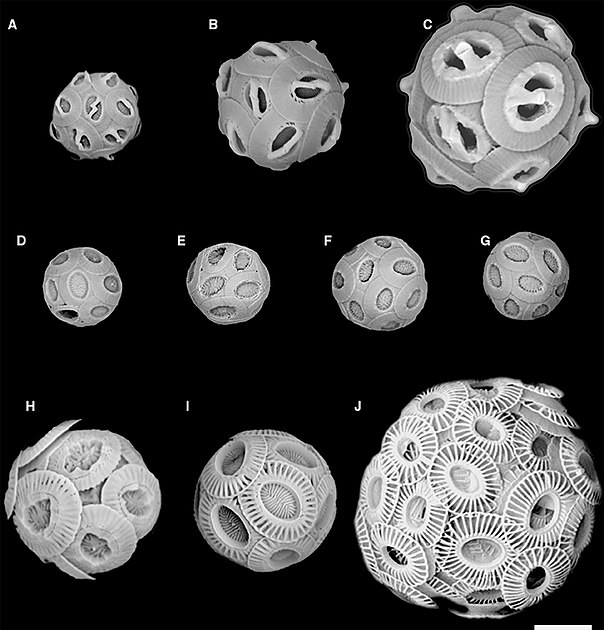
Scanning electron micrograph of Coccolithus pelagicus, plated with coccoliths
Coccoliths are individual plates or scales of calcium carbonate formed by coccolithophores (single-celled phytoplankton such as Emiliania huxleyi ) and cover the cell surface arranged in the form of a spherical shell, called a coccosphere .