Cutinase has an assigned enzyme commission number of EC 3.1.1.74.[2] Cutinase is in the third class of enzymes, meaning that its primary function is to hydrolyze its substrate (in this case, cutin).[3] Within the third class, cutinase is further categorized into the first subclass, which indicates that it specifically hydrolyzes ester bonds.[2] It is then placed in the first sub-subclass, meaning that it targets carboxylic esters, which are those that join together cutin polymers.[2]
Function
Cutin composes most of the waxy cuticle layer around plant cells. In order to enter plant cells, pathogens need to traverse this barrier.
Most plants have a layer composed of cutin, called the cuticle, on their aboveground surfaces such as stems, leaves, and fruits.[4] This layer of cutin is formed by a matrix-like structure that contains waxy components embedded in the carbohydrate layers.[5] The molecule, cutin, which composes most of the cuticle matrix (40-80%), is composed primarily of fatty acid chains that are polymerized via carboxylic ester bonds.[4][6]
Research suggests that cutin plays a critical role in preventing pathogenic infections in plant systems.[7] For instance, experiments conducted on tomato plants that had a substantial inability to synthesize cutin found that the tomatoes produced by those plants were significantly more susceptible to infection by both opportunistic pathogens and intentionally inoculated fungalspores.[8]
Cutinase is produced by a variety of fungal plant pathogens, and its activity was first detected in the fungus, Penicillium spinulosum.[9] In studies of Nectria haematococca, a fungal pathogen that is the cause of foot rot in pea plants, cutinase has been shown to play key roles in facilitating the early stages of plant infection.[9] It is also suggested that fungal spores that make initial contact with plant surfaces, a small amount of catalytic cutinase produces cutin monomers which in turn up-regulate the expression of the cutinase gene.[9] This proposes that the expression pathway of cutinase in fungal spores is characterized by a positive feedback loop until the fungus successfully breaches the cutin layer; however, the specific mechanism of this pathway is unclear.[9][10] Inhibition of cutinase has been shown to prevent fungal infection through intact cuticles.[10] Conversely, the supplementation of cutinase to fungi that are not able to produce it naturally had been shown to enhance fungal infection success rates.[9]
Cutinases have also been observed in a few plant pathogenic bacterial species, such as Streptomyces scabies, Thermobifida fusca, Pseudomonas mendocina, and Pseudomonas putida, but these have not been studied to the extent as those found in fungi.[11][12] The molecular structure of the Thermobifida fusca cutinase shows similarities to the Fusarium solani pisi fungal cutinase, with congruencies in their active sites and overall mechanisms.[11]
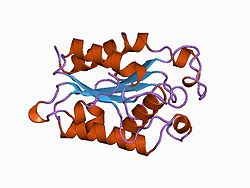
Structure
Cutinase belongs to the α-β class of proteins, with a central β-sheet of 5 parallel strands covered by 5 alpha helices on either side of the sheet.[13] Fungal cutinase is generally composed of around 197 amino acid residues, and its native form consists of a single domain.[14] The protein also contains 4 invariant cysteine residues that form 2 disulfide bridges, whose cleavage results in a complete loss of enzymatic activity.[15][14]
Crystal structures have shown that the active site of cutinases is found on one end of the ellipsoid shape of the enzyme.[16] This active site is seen flanked by two hydrophobic loop structures and partly covered by 2 thin bridges formed by amino acid side chains.[13][16] It does not possess a hydrophobic lid, which is a common constituent feature among other lipases.[13] Instead, the catalytic serine in the active site is exposed to open solvent, and the cutinase enzyme does not show interfacial activation behaviors at an aqueous-nonpolar interface.[13][14] Cutinase activation is believed to be derived from slight shifts in the conformation of hydrophobic residues, acting as a miniature lid.[13] The oxyanion hole in the active site is a constituent feature of the binding site, which differs from most lipolytic enzymes whose oxyanion holes are induced upon substrate binding.[17]
Mechanism
Cutinase is a serine esterase, and the active site contains a serine-histidine-aspartate triad and an oxyanion hole, which are signature elements of serine hydrolases.[15][18] The binding site of the cutin lipid polymer consists of two hydrophobic loops characterized by nonpolar amino acids such as leucine, alanine, isoleucine, and proline.[18] These hydrophobic residues show a higher degree of flexibility, suggesting an induced fit model to facilitate cutin bonding to the active site.[13] In the cutinase active site, histidine deprotonates serine, allowing the serine to undergo a nucleophilic attack on the cutin carboxylic ester.[19] This is followed by an elimination reaction whereby the charged oxygen (stabilized by the oxyanion hole) creates a double bond, removing an R group from the cutin polymer in the form of an alcohol.[19] The process repeats with a nucleophilic attack on the new carboxylic ester by a deprotonated water molecule.[19] Following this, the charged oxygen reforms its double bond, removing the serine attachment and releasing the carboxylic acid R monomer.[19]
Step by step mechanism of the hydrolysis of cutin polymers via the serine-histidine-aspartate residues in the active site of cutinase. Image adapted from Mei Leung, Gemma L. Holliday, and James Willey.
Applications
The stability of cutinases in higher temperatures (20-50°C) and its compatibility with other hydrolytic enzymes has potential applications in the detergent industry.[20] In fact, it has been shown that cutinases are more efficient at cleaving and eliminating non-calcium fats from clothing when compared against other industrial lipases.[21] Another advantage of cutinase in this industry is its ability to be catalytically active with both water- and lipid-soluble ester compounds, making it a more versatile degradative agent.[20] This versatility is also subjecting cutinase to experiments in enhancing the biofuel industry because of its ability to facilitate transesterification of biofuels in various solubility environments.[20]
Rather unexpectedly, the ability to degrade the cutin layer of plants and their fruits holds the potential to be beneficial to the fruit industry.[20] This is because the cuticle layer of fruits is a putative mechanism of water regulation, and the degradation of this layer subjects the fruits to water movement across its membrane.[22] By using cutinase to degrade the cuticle of fruits, industry makers can enhance the drying of fruits and more easily deliver preservatives and additives to the flesh of the fruit.[20]
↑ Longhi S, Czjzek M, Lamzin V, Nicolas A, Cambillau C (May 1997). "Atomic resolution (1.0 A) crystal structure of Fusarium solani cutinase: stereochemical analysis". Journal of Molecular Biology. 268 (4): 779–799. doi:10.1006/jmbi.1997.1000. PMID9175860.
1 2 Heredia A (March 2003). "Biophysical and biochemical characteristics of cutin, a plant barrier biopolymer". Biochimica et Biophysica Acta (BBA) - General Subjects. 1620 (1–3): 1–7. doi:10.1016/s0304-4165(02)00510-x. PMID12595066.
↑ Nicolas A, Egmond M, Verrips CT, de Vlieg J, Longhi S, Cambillau C, Martinez C (January 1996). "Contribution of cutinase serine 42 side chain to the stabilization of the oxyanion transition state". Biochemistry. 35 (2): 398–410. doi:10.1021/bi9515578. PMID8555209.
1 2 Martinez C, Nicolas A, van Tilbeurgh H, Egloff MP, Cudrey C, Verger R, Cambillau C (January 1994). "Cutinase, a lipolytic enzyme with a preformed oxyanion hole". Biochemistry. 33 (1): 83–89. doi:10.1021/bi00167a011. PMID8286366.
1 2 3 4 Leung M, Holliday G, Willey J. "Cutinase". Mechanism and Catalytic Site Atlas. Retrieved 2022-09-28.
↑ Egmond MR, van Bemmel CJ (1997). "[6] Impact of structural information on understanding lipolytic function". Impact of structural information on understanding lipolytic function. Methods in Enzymology. Vol.284. Elsevier. pp.119–129. doi:10.1016/s0076-6879(97)84008-6. ISBN9780121821852. PMID9379930.
↑ "5510131 Enzyme assisted degradation of surface membranes of harvested fruits and vegetables". Biotechnology Advances. 15 (1): 273. January 1997. doi:10.1016/s0734-9750(97)88551-5. ISSN0734-9750.
Shirke, Abhijit N.; White, Christine; Englaender, Jacob A.; Zwarycz, Allison; Butterfoss, Glenn L.; Linhardt, Robert J.; Gross, Richard A. (2018). "Stabilizing Leaf and Branch Compost Cutinase (LCC) with Glycosylation: Mechanism and Effect on PET Hydrolysis". Biochemistry. 57 (7): 1190–1200. doi:10.1021/acs.biochem.7b01189. PMID29328676.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.