Proteins are large biomolecules and macromolecules that comprise one or more long chains of amino acid residues. Proteins perform a vast array of functions within organisms, including catalysing metabolic reactions, DNA replication, responding to stimuli, providing structure to cells and organisms, and transporting molecules from one location to another. Proteins differ from one another primarily in their sequence of amino acids, which is dictated by the nucleotide sequence of their genes, and which usually results in protein folding into a specific 3D structure that determines its activity.

Pectin is a heteropolysaccharide, a structural acid contained in the primary lamella, in the middle lamella, and in the cell walls of terrestrial plants. The principal chemical component of pectin is galacturonic acid which was isolated and described by Henri Braconnot in 1825. Commercially produced pectin is a white-to-light-brown powder, produced from citrus fruits for use as an edible gelling agent, especially in jams and jellies, dessert fillings, medications, and sweets; and as a food stabiliser in fruit juices and milk drinks, and as a source of dietary fiber.
Pectinases are a group of enzymes that breaks down pectin, a polysaccharide found in plant cell walls, through hydrolysis, transelimination and deesterification reactions. Commonly referred to as pectic enzymes, they include pectolyase, pectozyme, and polygalacturonase, one of the most studied and widely used commercial pectinases. It is useful because pectin is the jelly-like matrix which helps cement plant cells together and in which other cell wall components, such as cellulose fibrils, are embedded. Therefore, pectinase enzymes are commonly used in processes involving the degradation of plant materials, such as speeding up the extraction of fruit juice from fruit, including apples and sapota. Pectinases have also been used in wine production since the 1960s. The function of pectinase in brewing is twofold, first it helps break down the plant material and so helps the extraction of flavors from the mash. Secondly the presence of pectin in finished wine causes a haze or slight cloudiness. Pectinase is used to break this down and so clear the wine.
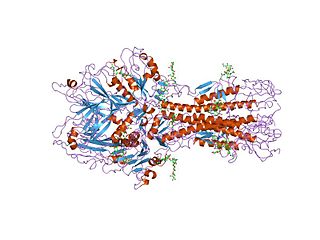
Hemagglutinin esterase (HEs) is a glycoprotein that certain enveloped viruses possess and use as an invading mechanism. HEs helps in the attachment and destruction of certain sialic acid receptors that are found on the host cell surface. Viruses that possess HEs include influenza C virus, toroviruses, and coronaviruses of the subgenus Embecovirus. HEs is a dimer transmembrane protein consisting of two monomers, each monomer is made of three domains. The three domains are: membrane fusion, esterase, and receptor binding domains.

A catalytic triad is a set of three coordinated amino acids that can be found in the active site of some enzymes. Catalytic triads are most commonly found in hydrolase and transferase enzymes. An acid-base-nucleophile triad is a common motif for generating a nucleophilic residue for covalent catalysis. The residues form a charge-relay network to polarise and activate the nucleophile, which attacks the substrate, forming a covalent intermediate which is then hydrolysed to release the product and regenerate free enzyme. The nucleophile is most commonly a serine or cysteine amino acid, but occasionally threonine or even selenocysteine. The 3D structure of the enzyme brings together the triad residues in a precise orientation, even though they may be far apart in the sequence.

Lipoxygenases (LOX) are a family of (non-heme) iron-containing enzymes, more specifically oxidative enzymes, most of which catalyze the dioxygenation of polyunsaturated fatty acids in lipids containing a cis,cis-1,4-pentadiene into cell signaling agents that serve diverse roles as autocrine signals that regulate the function of their parent cells, paracrine signals that regulate the function of nearby cells, and endocrine signals that regulate the function of distant cells.

Dickeya dadantii is a gram-negative bacillus that belongs to the family Pectobacteriaceae. It was formerly known as Erwinia chrysanthemi but was reassigned as Dickeya dadantii in 2005. Members of this family are facultative anaerobes, able to ferment sugars to lactic acid, have nitrate reductase, but lack oxidases. Even though many clinical pathogens are part of the order Enterobacterales, most members of this family are plant pathogens. D. dadantii is a motile, nonsporing, straight rod-shaped cell with rounded ends, much like the other members of the genus, Dickeya. Cells range in size from 0.8 to 3.2 μm by 0.5 to 0.8 μm and are surrounded by numerous flagella (peritrichous).
The pelB leader sequence is a sequence of amino acids which, when attached to a protein, directs the protein to the bacterial periplasm, where the sequence is removed by a signal peptidase. Specifically, pelB refers to pectate lyase B of Erwinia carotovora CE. The leader sequence consists of the 22 N-terminal amino acid residues. This leader sequence can be attached to any other protein resulting in a transfer of such a fused protein to the periplasmic space of Gram-negative bacteria, such as Escherichia coli, often used in genetic engineering. Protein secretion can increase the stability of cloned gene products. For instance it was shown that the half-life of the recombinant proinsulin is increased 10-fold when the protein is secreted to the periplasmic space.
Phospholipase D (EC 3.1.4.4, lipophosphodiesterase II, lecithinase D, choline phosphatase, PLD; systematic name phosphatidylcholine phosphatidohydrolase) is an enzyme of the phospholipase superfamily that catalyses the following reaction

PDE3 is a phosphodiesterase. The PDEs belong to at least eleven related gene families, which are different in their primary structure, substrate affinity, responses to effectors, and regulation mechanism. Most of the PDE families are composed of more than one gene. PDE3 is clinically significant because of its role in regulating heart muscle, vascular smooth muscle and platelet aggregation. PDE3 inhibitors have been developed as pharmaceuticals, but their use is limited by arrhythmic effects and they can increase mortality in some applications.

Sucrase-isomaltase is a bifunctional glucosidase located on the brush border of the small intestine, encoded by the human gene SI. It is a dual-function enzyme with two GH31 domains, one serving as the isomaltase, the other as a sucrose alpha-glucosidase. It has preferential expression in the apical membranes of enterocytes. The enzyme’s purpose is to digest dietary carbohydrates such as starch, sucrose and isomaltose. By further processing the broken-down products, energy in the form of ATP can be generated.

Chlorophyllase is an essential enzyme in chlorophyll metabolism. It is a membrane proteins commonly known as chlase (EC 3.1.1.14, CLH) with systematic name chlorophyll chlorophyllidohydrolase. It catalyzes the reaction
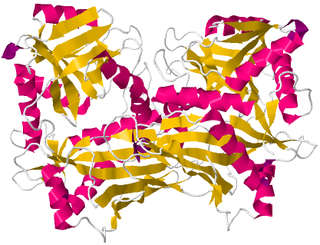
Branched-chain amino acid aminotransferase (BCAT), also known as branched-chain amino acid transaminase, is an aminotransferase enzyme (EC 2.6.1.42) which acts upon branched-chain amino acids (BCAAs). It is encoded by the BCAT2 gene in humans. The BCAT enzyme catalyzes the conversion of BCAAs and α-ketoglutarate into branched chain α-keto acids and glutamate.

Tyrosylprotein sulfotransferase is an enzyme that catalyzes tyrosine sulfation.

Wall-associated kinases (WAKs) are one of many classes of plant proteins known to serve as a medium between the extracellular matrix (ECM) and cytoplasm of cell walls. They are serine-threonine kinases that contain epidermal growth factor (EGF) repeats, a cytoplasmic kinase and are located in the cell walls. They provide a linkage between the inner and outer surroundings of cell walls. WAKs are under a group of receptor-like kinases (RLK) that are actively involved in sensory and signal transduction pathways especially in response to foreign attacks by pathogens and in cell development. On the other hand, pectins are an abundant group of complex carbohydrates present in the primary cell wall that play roles in cell growth and development, protection, plant structure and water holding capacity.
Pectate lyase is an enzyme involved in the maceration and soft rotting of plant tissue. Pectate lyase is responsible for the eliminative cleavage of pectate, yielding oligosaccharides with 4-deoxy-α-D-mann-4-enuronosyl groups at their non-reducing ends. The protein is maximally expressed late in pollen development. It has been suggested that the pollen expression of pectate lyase genes might relate to a requirement for pectin degradation during pollen tube growth.
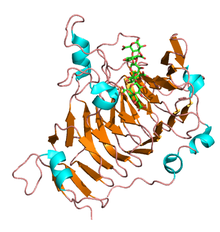
Pectin lyase is a polysaccharide enzyme with a complex structure that is present in plant cell walls. It has a significant role in pectin degradation and different biotechnological and industrial applications. It can be found in different organisms.
The enzyme tannase (EC 3.1.1.20) catalyzes the following reaction:
Reticulons are a group of evolutionary conservative proteins residing predominantly in endoplasmic reticulum, primarily playing a role in promoting membrane curvature. In addition, reticulons may play a role in nuclear pore complex formation, vesicle formation, and other processes yet to be defined. They have also been linked to oligodendrocyte roles in inhibition of neurite outgrowth. Some studies link RTNs with Alzheimer's disease and amyotrophic lateral sclerosis.

Endo-polygalacturonase (EC 3.2.1.15, pectin depolymerase, pectolase, pectin hydrolase, and poly-α-1,4-galacturonide glycanohydrolase; systematic name (1→4)-α-D-galacturonan glycanohydrolase (endo-cleaving)) is an enzyme that hydrolyzes the α-1,4 glycosidic bonds between galacturonic acid residues: