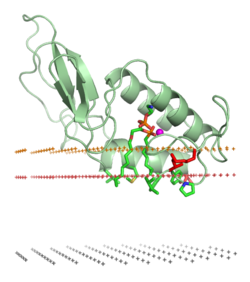
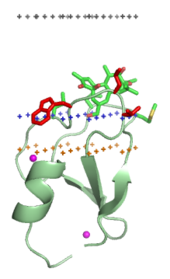
Peripheral membrane proteins, or extrinsic membrane proteins, [1] are membrane proteins that adhere only temporarily to the biological membrane with which they are associated. These proteins attach to integral membrane proteins, or penetrate the peripheral regions of the lipid bilayer. The regulatory protein subunits of many ion channels and transmembrane receptors, for example, may be defined as peripheral membrane proteins. In contrast to integral membrane proteins, peripheral membrane proteins tend to collect in the water-soluble component, or fraction, of all the proteins extracted during a protein purification procedure. Proteins with GPI anchors are an exception to this rule and can have purification properties similar to those of integral membrane proteins.
Contents
- Binding to the lipid bilayer
- Membrane binding mechanisms
- Non-specific hydrophobic association
- Covalently bound lipid anchors
- Specific protein–lipid binding
- Protein–lipid electrostatic interactions
- Spatial position in membrane
- Categories
- Enzymes
- Membrane-targeting domains ("lipid clamps")
- Structural domains
- Transporters of small hydrophobic molecules
- Electron carriers
- Polypeptide hormones, toxins, and antimicrobial peptides
- See also
- References
- Further reading
- External links
The reversible attachment of proteins to biological membranes has shown to regulate cell signaling and many other important cellular events, through a variety of mechanisms. [2] For example, the close association between many enzymes and biological membranes may bring them into close proximity with their lipid substrate(s). [3] Membrane binding may also promote rearrangement, dissociation, or conformational changes within many protein structural domains, resulting in an activation of their biological activity. [4] [5] Additionally, the positioning of many proteins are localized to either the inner or outer surfaces or leaflets of their resident membrane. [6] This facilitates the assembly of multi-protein complexes by increasing the probability of any appropriate protein–protein interactions.