
In bacteriology, gram-positive bacteria are bacteria that give a positive result in the Gram stain test, which is traditionally used to quickly classify bacteria into two broad categories according to their type of cell wall.

Gram-negative bacteria are bacteria that do not retain the crystal violet stain used in the Gram staining method of bacterial differentiation. They are characterized by their cell envelopes, which are composed of a thin peptidoglycan cell wall sandwiched between an inner cytoplasmic cell membrane and a bacterial outer membrane.
In cell biology, an organelle is a specialized subunit, usually within a cell, that has a specific function. The name organelle comes from the idea that these structures are parts of cells, as organs are to the body, hence organelle, the suffix -elle being a diminutive. Organelles are either separately enclosed within their own lipid bilayers or are spatially distinct functional units without a surrounding lipid bilayer. Although most organelles are functional units within cells, some functional units that extend outside of cells are often termed organelles, such as cilia, the flagellum and archaellum, and the trichocyst.

The phylum "Bacteroidetes" is composed of three large classes of Gram-negative, nonsporeforming, anaerobic or aerobic, and rod-shaped bacteria that are widely distributed in the environment, including in soil, sediments, and sea water, as well as in the guts and on the skin of animals.
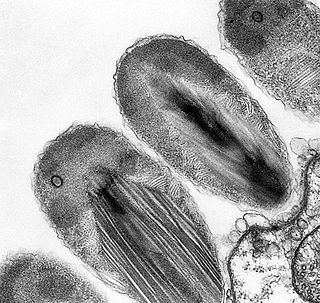
Verrucomicrobia is a phylum of Gram-negative bacteria that contains only a few described species. The species identified have been isolated from fresh water, marine and soil environments and human faeces. A number of as-yet uncultivated species have been identified in association with eukaryotic hosts including extrusive explosive ectosymbionts of protists and endosymbionts of nematodes residing in their gametes.

The Chlamydiae are a bacterial phylum and class whose members are remarkably diverse, including pathogens of humans and animals, symbionts of ubiquitous protozoa, and marine sediment forms not yet well understood. All of the Chlamydiae that humans have known about for many decades are obligate intracellular bacteria; in 2020 many additional Chlamydiae were discovered in ocean-floor environments, and it is not yet known whether they all have hosts. Historically it was believed that all Chlamydiae had a peptidoglycan-free cell wall, but studies in the 2010s demonstrated a detectable presence of peptidoglycan, as well as other important proteins.

The periplasm is a concentrated gel-like matrix in the space between the inner cytoplasmic membrane and the bacterial outer membrane called the periplasmic space in gram-negative bacteria. Using cryo-electron microscopy it has been found that a much smaller periplasmic space is also present in gram-positive bacteria.

Anammox, an abbreviation for anaerobic ammonium oxidation, is a globally important microbial process of the nitrogen cycle that takes place in many natural environments. The bacteria mediating this process were identified in 1999, and were a great surprise for the scientific community. In the anammox reaction, nitrite and ammonium ions are converted directly into diatomic nitrogen and water.
The bacterium, despite its simplicity, contains a well-developed cell structure which is responsible for some of its unique biological structures and pathogenicity. Many structural features are unique to bacteria and are not found among archaea or eukaryotes. Because of the simplicity of bacteria relative to larger organisms and the ease with which they can be manipulated experimentally, the cell structure of bacteria has been well studied, revealing many biochemical principles that have been subsequently applied to other organisms.

An alpha solenoid is a protein fold composed of repeating alpha helix subunits, commonly helix-turn-helix motifs, arranged in antiparallel fashion to form a superhelix. Alpha solenoids are known for their flexibility and plasticity. Like beta propellers, alpha solenoids are a form of solenoid protein domain commonly found in the proteins comprising the nuclear pore complex. They are also common in membrane coat proteins known as coatomers, such as clathrin, and in regulatory proteins that form extensive protein-protein interactions with their binding partners. Examples of alpha solenoid structures binding RNA and lipids have also been described.

Gracilicutes is a clade in bacterial phylogeny.
The PVC superphylum is a superphylum of bacteria named after its three important members, Planctomycetes, Verrucomicrobia, and Chlamydiae. Cavalier-Smith postulated that the PVC bacteria probably lost or reduced their peptidoglycan cell wall twice. It has been hypothesised that a member of the PVC clade might have been the host cell in the endosymbiotic event that gave rise to the first proto-eukaryotic cell.
Epixenosomes, also known as Candidatus Epixenosoma are a genus of bacteria in the phylum Verrucomicrobia that form a symbiosis with marine ciliates of the genus Euplotidium, where they help to defend their ciliate hosts against predators. It is a monospecific genus, containing only the species Ca. Epixenosoma ejectans.
The FCB group is a superphylum of bacteria named after the main member phyla Fibrobacteres, Chlorobi, and Bacteroidetes. The members are considered to form a clade due to a number of conserved signature indels.
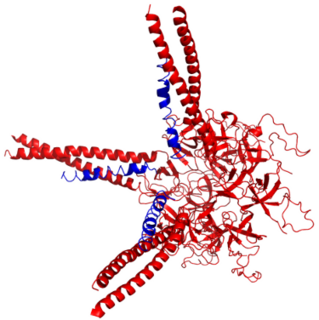
Prokaryotic ubiquitin-like protein (Pup) is a functional analog of ubiquitin found in the prokaryote Mycobacterium tuberculosis. Like ubiquitin, Pup serves to direct proteins to the proteasome for degradation in the Pup-proteasome system (PPS). However, the enzymology of ubiquitylation and pupylation is different, owing to their distinct evolutionary origins. In contrast to the three-step reaction of ubiquitylation, pupylation requires only two steps, and thus only two enzymes are involved in pupylation. The enzymes involved in pupylation are descended from glutamine synthetase.
The eocyte hypothesis is a biological classification that indicates eukaryotes emerged within the prokaryotic Crenarchaeota, a phylum within the archaea. This hypothesis was originally proposed by James A. Lake and colleagues in 1984 based on the discovery that the shapes of ribosomes in the Crenarchaeota and eukaryotes are more similar to each other than to either bacteria or the second major phylum of archaea, the Euryarchaeota.
"Candidatus Scalindua" is a bacterial genus, and a proposed member of the order Planctomycetes. These bacteria lack peptidoglycan in their cell wall and have a compartmentalized cytoplasm. They are ammonium oxidizing bacteria found in marine environments.
Poribacteria are a candidate phylum of bacteria originally discovered in the microbiome of marine sponges (Porifera). Poribacteria are Gram-negative primarily aerobic mixotrophs with the ability for oxidative phosphorylation, glycolysis, and autotrophic carbon fixation via the Wood – Ljungdahl pathway. Poribacterial heterotrophy is characterised by an enriched set of glycoside hydrolases, uronic acid degradation, as well as several specific sulfatases. This heterotrophic repertoire of poribacteria was suggested to be involved in the degradation of the extracellular sponge host matrix.
Lokiarchaeota is a proposed phylum of the Archaea. The phylum includes all members of the group previously named Deep Sea Archaeal Group (DSAG), also known as Marine Benthic Group B (MBG-B). A phylogenetic analysis disclosed a monophyletic grouping of the Lokiarchaeota with the eukaryotes. The analysis revealed several genes with cell membrane-related functions. The presence of such genes support the hypothesis of an archaeal host for the emergence of the eukaryotes; the eocyte-like scenarios.
Gemmata obscuriglobus is a species of Gram-negative, aerobic, heterotrophic bacteria of the phylum Planctomycetes. G. obscuriglobus occur in freshwater habitats and was first described in 1984, and is the only described species in its genus.







