
Autophagy is the natural, conserved degradation of the cell that removes unnecessary or dysfunctional components through a lysosome-dependent regulated mechanism. It allows the orderly degradation and recycling of cellular components. Although initially characterized as a primordial degradation pathway induced to protect against starvation, it has become increasingly clear that autophagy also plays a major role in the homeostasis of non-starved cells. Defects in autophagy have been linked to various human diseases, including neurodegeneration and cancer, and interest in modulating autophagy as a potential treatment for these diseases has grown rapidly.
Autophagin-1 (Atg4/Apg4) is a unique cysteine protease responsible for the cleavage of the carboxyl terminus of Atg8/Apg8/Aut7, a reaction essential for its lipidation during autophagy. Human Atg4 homologues cleave the carboxyl termini of the three human Atg8 homologues, microtubule-associated protein light chain 3 (LC3), GABARAP, and GATE-16.

Vojo Deretic, is distinguished professor and chair of the Department of Molecular Genetics and Microbiology at the University of New Mexico School of Medicine. Deretic was the founding director of the Autophagy, Inflammation and Metabolism (AIM) Center of Biomedical Research Excellence. The AIM center promotes autophagy research nationally and internationally.

Autophagy protein 5 (ATG5) is a protein that, in humans, is encoded by the ATG5 gene located on chromosome 6. It is an E3 ubi autophagic cell death. ATG5 is a key protein involved in the extension of the phagophoric membrane in autophagic vesicles. It is activated by ATG7 and forms a complex with ATG12 and ATG16L1. This complex is necessary for LC3-I conjugation to PE (phosphatidylethanolamine) to form LC3-II. ATG5 can also act as a pro-apoptotic molecule targeted to the mitochondria. Under low levels of DNA damage, ATG5 can translocate to the nucleus and interact with survivin.

Microtubule-associated proteins 1A/1B light chain 3B is a protein that in humans is encoded by the MAP1LC3B gene. LC3 is a central protein in the autophagy pathway where it functions in autophagy substrate selection and autophagosome biogenesis. LC3 is the most widely used marker of autophagosomes.

Microtubule-associated proteins 1A/1B light chain 3A is a protein that in humans is encoded by the MAP1LC3A gene. Two transcript variants encoding different isoforms have been found for this gene.

Cysteine protease ATG4B is an enzyme that in humans is encoded by the ATG4B gene.

Autophagy related 16 like 1 is a protein that in humans is encoded by the ATG16L1 gene. This protein is characterized as a subunit of the autophagy-related ATG12-ATG5/ATG16 complex and is essentially important for the LC3 (ATG8) lipidation and autophagosome formation. This complex localizes to the membrane and is released just before or after autophagosome completion.

Autophagy related 12 is a protein that in humans is encoded by the ATG12 gene.

Autophagy related 7 is a protein in humans encoded by ATG7 gene. Related to GSA7; APG7L; APG7-LIKE.
AuTophaGy related 1 (Atg1) is a 101.7kDa serine/threonine kinase in S.cerevisiae, encoded by the gene ATG1. It is essential for the initial building of the autophagosome and Cvt vesicles. In a non-kinase role it is - through complex formation with Atg13 and Atg17 - directly controlled by the TOR kinase, a sensor for nutrient availability.
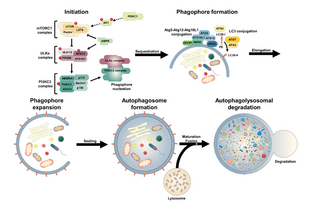
An autophagosome is a spherical structure with double layer membranes. It is the key structure in macroautophagy, the intracellular degradation system for cytoplasmic contents. After formation, autophagosomes deliver cytoplasmic components to the lysosomes. The outer membrane of an autophagosome fuses with a lysosome to form an autolysosome. The lysosome's hydrolases degrade the autophagosome-delivered contents and its inner membrane.
In molecular biology, autophagy related 3 (Atg3) is the E2 enzyme for the LC3 lipidation process. It is essential for autophagy. The super protein complex, the Atg16L complex, consists of multiple Atg12-Atg5 conjugates. Atg16L has an E3-like role in the LC3 lipidation reaction. The activated intermediate, LC3-Atg3 (E2), is recruited to the site where the lipidation takes place.

Yoshinori Ohsumi is a Japanese cell biologist specializing in autophagy, the process that cells use to destroy and recycle cellular components. Ohsumi is a professor at Tokyo Institute of Technology's Institute of Innovative Research. He received the Kyoto Prize for Basic Sciences in 2012, the 2016 Nobel Prize in Physiology or Medicine, and the 2017 Breakthrough Prize in Life Sciences for his discoveries of mechanisms for autophagy.
Chaperone-assisted selective autophagy is a cellular process for the selective, ubiquitin-dependent degradation of chaperone-bound proteins in lysosomes.

Omegasome is a cell organelle consisting of lipid bilayer membranes enriched for phosphatidylinositol 3-phosphate, and related to a process of autophagy. It is a subdomain of the Endoplasmic Reticulum (ER), and has a morphology resembling the Greek capital letter Omega (Ω). Omegasomes are the sites from which phagophores form, which are sack-like structures that mature into autophagosomes, and fuse with lysosomes in order to degrade the contents of the autophagosomes. The formation of omegasomes depends on various factors, however in general, formation of omegasomes is increased as a response to starvation, and in some biochemical situations the presence of PI(3)P leads to the formation of omegasomes.
Microautophagy is one of the three common forms of autophagic pathway, but unlike macroautophagy and chaperone-mediated autophagy, it is mediated—in mammals by lysosomal action or in plants and fungi by vacuolar action—by direct engulfment of the cytoplasmic cargo. Cytoplasmic material is trapped in the lysosome/vacuole by a random process of membrane invagination.
Rab GTPases are molecular switches that regulate membrane traffic. They are active in their GTP-bound form and inactive when bound to GDP. The GTPase YPT1, and its mammalian homologue Rab1, regulate membrane-tethering events on three different pathways: autophagy, ER-Golgi, and intra-Golgi traffic. In the yeast Saccharomyces cerevisiae, many of the ATG proteins needed for macroautophagy are shared with the biosynthetic cytoplasm to the vacuole-targeting (CVT) pathway that transports certain hydrolases into the vacuole. Both pathways require YPT1; however, only the macroautophagy pathway is conserved in higher eukaryotes. In the macroautophagy pathway, Rab1 mediates the recruitment of Atg1 to the PAS. Rab1 regulates macroautophagy by recruiting its effector, Atg1, to the PAS to tether Atg9 vesicles to each other or to other membranes.

Ubiquitin-like proteins (UBLs) are a family of small proteins involved in post-translational modification of other proteins in a cell, usually with a regulatory function. The UBL protein family derives its name from the first member of the class to be discovered, ubiquitin (Ub), best known for its role in regulating protein degradation through covalent modification of other proteins. Following the discovery of ubiquitin, many additional evolutionarily related members of the group were described, involving parallel regulatory processes and similar chemistry. UBLs are involved in a widely varying array of cellular functions including autophagy, protein trafficking, inflammation and immune responses, transcription, DNA repair, RNA splicing, and cellular differentiation.
