
Proteasomes are protein complexes which degrade unneeded or damaged proteins by proteolysis, a chemical reaction that breaks peptide bonds. Enzymes that help such reactions are called proteases.

Ubiquitin is a small regulatory protein found in most tissues of eukaryotic organisms, i.e., it is found ubiquitously. It was discovered in 1975 by Gideon Goldstein and further characterized throughout the late 1970s and 1980s. Four genes in the human genome code for ubiquitin: UBB, UBC, UBA52 and RPS27A.

A ubiquitin ligase is a protein that recruits an E2 ubiquitin-conjugating enzyme that has been loaded with ubiquitin, recognizes a protein substrate, and assists or directly catalyzes the transfer of ubiquitin from the E2 to the protein substrate. In simple and more general terms, the ligase enables movement of ubiquitin from a ubiquitin carrier to another thing by some mechanism. The ubiquitin, once it reaches its destination, ends up being attached by an isopeptide bond to a lysine residue, which is part of the target protein. E3 ligases interact with both the target protein and the E2 enzyme, and so impart substrate specificity to the E2. Commonly, E3s polyubiquitinate their substrate with Lys48-linked chains of ubiquitin, targeting the substrate for destruction by the proteasome. However, many other types of linkages are possible and alter a protein's activity, interactions, or localization. Ubiquitination by E3 ligases regulates diverse areas such as cell trafficking, DNA repair, and signaling and is of profound importance in cell biology. E3 ligases are also key players in cell cycle control, mediating the degradation of cyclins, as well as cyclin dependent kinase inhibitor proteins. The human genome encodes over 600 putative E3 ligases, allowing for tremendous diversity in substrates.

Parkin is a 465-amino acid residue E3 ubiquitin ligase, a protein that in humans and mice is encoded by the PARK2 gene. Parkin plays a critical role in ubiquitination – the process whereby molecules are covalently labelled with ubiquitin (Ub) and directed towards degradation in proteasomes or lysosomes. Ubiquitination involves the sequential action of three enzymes. First, an E1 ubiquitin-activating enzyme binds to inactive Ub in eukaryotic cells via a thioester bond and mobilises it in an ATP-dependent process. Ub is then transferred to an E2 ubiquitin-conjugating enzyme before being conjugated to the target protein via an E3 ubiquitin ligase. There exists a multitude of E3 ligases, which differ in structure and substrate specificity to allow selective targeting of proteins to intracellular degradation.

Ubiquitin-like modifier activating enzyme 1 (UBA1) is an enzyme which in humans is encoded by the UBA1 gene. UBA1 participates in ubiquitination and the NEDD8 pathway for protein folding and degradation, among many other biological processes. This protein has been linked to X-linked spinal muscular atrophy type 2, neurodegenerative diseases, and cancers.

Deubiquitinating enzymes (DUBs), also known as deubiquitinating peptidases, deubiquitinating isopeptidases, deubiquitinases, ubiquitin proteases, ubiquitin hydrolases, or ubiquitin isopeptidases, are a large group of proteases that cleave ubiquitin from proteins. Ubiquitin is attached to proteins in order to regulate the degradation of proteins via the proteasome and lysosome; coordinate the cellular localisation of proteins; activate and inactivate proteins; and modulate protein-protein interactions. DUBs can reverse these effects by cleaving the peptide or isopeptide bond between ubiquitin and its substrate protein. In humans there are nearly 100 DUB genes, which can be classified into two main classes: cysteine proteases and metalloproteases. The cysteine proteases comprise ubiquitin-specific proteases (USPs), ubiquitin C-terminal hydrolases (UCHs), Machado-Josephin domain proteases (MJDs) and ovarian tumour proteases (OTU). The metalloprotease group contains only the Jab1/Mov34/Mpr1 Pad1 N-terminal+ (MPN+) (JAMM) domain proteases.
Chemical modification refers to a number of various processes involving the alteration of the chemical constitution or structure of molecules.

Endoplasmic-reticulum-associated protein degradation (ERAD) designates a cellular pathway which targets misfolded proteins of the endoplasmic reticulum for ubiquitination and subsequent degradation by a protein-degrading complex, called the proteasome.
Ubiquitin-conjugating enzymes, also known as E2 enzymes and more rarely as ubiquitin-carrier enzymes, perform the second step in the ubiquitination reaction that targets a protein for degradation via the proteasome. The ubiquitination process covalently attaches ubiquitin, a short protein of 76 amino acids, to a lysine residue on the target protein. Once a protein has been tagged with one ubiquitin molecule, additional rounds of ubiquitination form a polyubiquitin chain that is recognized by the proteasome's 19S regulatory particle, triggering the ATP-dependent unfolding of the target protein that allows passage into the proteasome's 20S core particle, where proteases degrade the target into short peptide fragments for recycling by the cell.

CDC34 is a gene that in humans encodes the protein Ubiquitin-conjugating enzyme E2 R1. This protein is a member of the ubiquitin-conjugating enzyme family, which catalyzes the covalent attachment of ubiquitin to other proteins.

Ubiquitin D is a protein that in humans is encoded by the UBD gene, also known as FAT10. UBD acts like ubiquitin, by covalently modifying proteins and tagging them for destruction in the proteasome.

NEDD8-activating enzyme E1 regulatory subunit is a protein that in humans is encoded by the NAE1 gene.

NEDD8-activating enzyme E1 catalytic subunit is a protein that in humans is encoded by the UBA3 gene.

26S proteasome non-ATPase regulatory subunit 14, also known as 26S proteasome non-ATPase subunit Rpn11, is an enzyme that in humans is encoded by the PSMD14 gene. This protein is one of the 19 essential subunits of the complete assembled 19S proteasome complex. Nine subunits Rpn3, Rpn5, Rpn6, Rpn7, Rpn8, Rpn9, Rpn11, SEM1, and Rpn12 form the lid sub complex of the 19S regulatory particle of the proteasome complex.

Autophagy related 7 is a protein in humans encoded by ATG7 gene. Related to GSA7; APG7L; APG7-LIKE.

Pyruvate dehydrogenase (lipoamide) beta, also known as pyruvate dehydrogenase E1 component subunit beta, mitochondrial or PDHE1-B is an enzyme that in humans is encoded by the PDHB gene. The pyruvate dehydrogenase (PDH) complex is a nuclear-encoded mitochondrial multienzyme complex that catalyzes the overall conversion of pyruvate to acetyl-CoA and CO2, and provides the primary link between glycolysis and the tricarboxylic acid (TCA) cycle. The PDH complex is composed of multiple copies of three enzymatic components: pyruvate dehydrogenase (E1), dihydrolipoamide acetyltransferase (E2) and lipoamide dehydrogenase (E3). The E1 enzyme is a heterotetramer of two alpha and two beta subunits. This gene encodes the E1 beta subunit. Mutations in this gene are associated with pyruvate dehydrogenase E1-beta deficiency.
Ubiquitin-like 1-activating enzyme E1B (UBLE1B) also known as SUMO-activating enzyme subunit 2 (SAE2) is an enzyme that in humans is encoded by the UBA2 gene.
A proteolysis targeting chimera (PROTAC) is a heterobifunctional molecule composed of two active domains and a linker, capable of removing specific unwanted proteins. Rather than acting as a conventional enzyme inhibitor, a PROTAC works by inducing selective intracellular proteolysis. PROTACs consist of two covalently linked protein-binding molecules: one capable of engaging an E3 ubiquitin ligase, and another that binds to a target protein meant for degradation. Recruitment of the E3 ligase to the target protein results in ubiquitination and subsequent degradation of the target protein via the proteasome. Because PROTACs need only to bind their targets with high selectivity, there are currently many efforts to retool previously ineffective inhibitor molecules as PROTACs for next-generation drugs.

Ubiquitin conjugating enzyme E2 Z (UBE2Z), also known as UBA6-specific E2 enzyme 1 (USE1), is an enzyme that in humans is encoded by the UBE2Z gene on chromosome 17. It is ubiquitously expressed in many tissues and cell types. UBE2Z is an E2 ubiquitin conjugating enzyme and participates in the second step of protein ubiquitination during proteolysis. A genome-wide association study (GWAS) revealed the UBE2Z gene to be associated with chronic kidney disease. The UBE2Z gene also contains one of 27 SNPs associated with increased risk of coronary artery disease.

The ubiquitin carboxyl-terminal hydrolase 27, also known as deubiquitinating enzyme 27, ubiquitin thioesterase 27 and USP27X, is a deubiquitinating enzyme which is mainly characterized for cleaving ubiquitin (Ub) from proteins and other molecules. Ubiquitin binds to proteins so as to regulate the degradation of them via the proteasome and lysosome among many other functions.
 Figure 1. shows the sequence of how the ubiquitin activating enzyme attaches to ATP and the ubiquitin substrate. It also shows how two ubiquitin substrates can be bound at one time. [6]
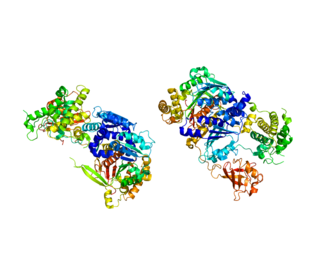
Figure 1. shows the sequence of how the ubiquitin activating enzyme attaches to ATP and the ubiquitin substrate. It also shows how two ubiquitin substrates can be bound at one time. [6] Figure 2. E1 protein binds a molecule of ubiquitin in each of two identical active sites (highlighted). The important residues, Cysteine and Arginine, are labeled in red. [2]
Figure 2. E1 protein binds a molecule of ubiquitin in each of two identical active sites (highlighted). The important residues, Cysteine and Arginine, are labeled in red. [2] Figure 3. Close-up view of the unbound active site. Arg (603) is believed to recharge the catalytic Cys (600) once ubiquitin has been transferred to the E2 enzyme. [2]
Figure 3. Close-up view of the unbound active site. Arg (603) is believed to recharge the catalytic Cys (600) once ubiquitin has been transferred to the E2 enzyme. [2]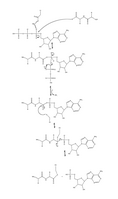 Figure 4. Full mechanism for adenylation of ubiquitin and subsequent ubiquitin binding to E1.
Figure 4. Full mechanism for adenylation of ubiquitin and subsequent ubiquitin binding to E1.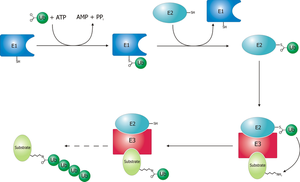 Schematic diagram of the ubiquitylation system.
Schematic diagram of the ubiquitylation system.




