Adenine phosphoribosyltransferase (APRTase) is an enzyme encoded by the APRTgene, found in humans on chromosome 16.[5] It is part of the Type I PRTase family and is involved in the nucleotide salvage pathway, which provides an alternative to nucleotide biosynthesis de novo in humans and most other animals.[6] In parasitic protozoa such as giardia, APRTase provides the sole mechanism by which AMP can be produced.[7] APRTase deficiency contributes to the formation of kidney stones (urolithiasis) and to potential kidney failure.[8]
The APRT gene is constituted by 5 exons (in blue). The start (ATG) and stop (TGA) codons are indicated (bold blue). CpG dinucleotides are emphasized in red. They are more abundant in the upstream region of the gene where they form a CpG island.
Function
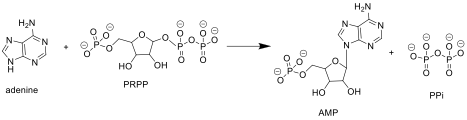
APRTase catalyzes the following reaction in the purine nucleotide salvage pathway:
ARPTase catalyzes a phosphoribosyl transfer from PRPP to adenine, forming AMP and releasing pyrophosphate (PPi).
In organisms that can synthesize purines de novo, the nucleotide salvage pathway provides an alternative that is energetically more efficient. It can salvage adenine from the polyamine biosynthetic pathway or from dietary sources of purines.[6] Although APRTase is functionally redundant in these organisms, it becomes more important during periods of rapid growth, such as embryogenesis and tumor growth.[9] It is constitutively expressed in all mammalian tissue.[10]
In protozoan parasites, the nucleotide salvage pathway provides the sole means for nucleotide synthesis. Since the consequences of APRTase deficiency in humans is comparatively mild and treatable, it may be possible to treat certain parasitic infections by targeting APRTase function.[11]
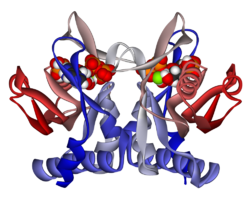
APRTase is a homodimer, with 179 amino acid residues per monomer. Each monomer contains the following regions:
Catalytic site of APRTase with reactants adenine and PRPP resolved. The Hood is believed to be important for purine specificity, while the flexible loop is thought to contain the molecules within the active site.
"Core" domain (residues 33-169) with five parallel β-sheets
"Hood" domain (residues 5-34) with 2 α-helices and 2 β-sheets
"Flexible loop" domain (residues 95-113) with 2 antiparallel β-sheets[10]
Residues A131, L159, V25, and R27 are important for purine specificity in human APRTase.
The core is highly conserved across many PRTases. The hood, which contains the adeninebinding site, has more variability within the family of enzymes. A 13-residue motif comprises the PRPP binding region and involves two adjacent acidic residues and at least one surrounding hydrophobic residue.[13]
The enzyme's specificity for adenine involves hydrophobic residues Ala131 and Leu159 in the core domain. In humans, two residues in the hood domain hydrogen bond with the purine for further specificity: Val25 with the hydrogens on N6, and Arg27 with N1. Although the flexible loop does not interact with the hood during purine recognition, it is thought to close over the active site and sequester the reaction from solvents.[10]
Most research on APRTase reports that Mg2+ is essential for phosphoribosyl transfer, and this is conserved across Type I PRTases.[12] However, a recent effort to resolve the structure of human APRTase was unable to locate a single site for Mg2+, but did find evidence to suggest a Cl− atom near Trp98. Despite the difficulty of placing Mg2+, it is generally accepted that the catalytic mechanism is dependent on this ion.[6]
Mechanism
APRTase proceeds via a bi bi ordered sequential mechanism, involving the formation of a ternary complex. The enzyme first binds PRPP, followed by adenine. After the phosphoribosyl transfer occurs, pyrophosphate leaves first, followed by AMP. Kinetic studies indicate that the phosphoribosyl transfer is relatively fast, while the product release (particularly the release of AMP) is rate-limiting.[9]
In human APRTase, it is thought that adenine's N9 proton is abstracted by Glu104 to form an oxacarbenium transition state. This functions as the nucleophile to attack the anomeric carbon of PRPP, forming AMP and displacing pyrophosphate from PRPP. The mechanism of APRTase is generally consistent with that of other PRTases, which conserve the function of displacing PRPP's α-1-pyrophosphate using a nitrogen nucleophile, in either an SN1 or SN2 attack.[6]
ARPTase deficiency was first diagnosed in the UK in 1976. Since then, two categories of APRTase deficiency have been defined in humans.[14]
Type I deficiency results in a complete loss of APRTase activity and can occur in patients that are homozygous or compound heterozygous for various mutations.[15]Sequencing has revealed many different mutations that can account for Type 1, including missense mutations, nonsense mutations, a duplicated set of 4 base pairs in exon 3,[16] and a single thymineinsertion in intron 4.[17] These mutations cause effects that are clustered into three main areas: in the binding of PRPP's β-phosphate, in the binding of PRPP's 5'-phosphate, and in the segment of the flexible loop that closes over the active site during catalysis [10] Type I deficiency has been observed in various ethnic groups but studied predominately among White populations.[17]
Type II deficiency causes APRTase to have a reduced affinity for PRPP, resulting in a tenfold increase in the KM value.[6] It has been observed and studied primarily in Japan.[17]
A diagnosis of APRTase deficiency can be made by analyzing kidney stones, measuring DHA concentrations in urine, or analyzing APRTase activity in erythrocytes. It is treatable with regular doses of allopurinol or febuxostat, which inhibit xanthine dehydrogenase activity to prevent the accumulation and precipitation of DHA.[18] The condition can also be attenuated with a low-purine diet and high fluid intake.[14]
12Shi W, Tanaka KS, Crother TR, Taylor MW, Almo SC, Schramm VL (Sep 2001). "Structural analysis of adenine phosphoribosyltransferase from Saccharomyces cerevisiae". Biochemistry. 40 (36): 10800–9. doi:10.1021/bi010465h. PMID11535055.
12Bashor C, Denu JM, Brennan RG, Ullman B (Mar 2002). "Kinetic mechanism of adenine phosphoribosyltransferase from Leishmania donovani". Biochemistry. 41 (12): 4020–31. doi:10.1021/bi0158730. PMID11900545.
1234Silva M, Silva CH, Iulek J, Thiemann OH (Jun 2004). "Three-dimensional structure of human adenine phosphoribosyltransferase and its relation to DHA-urolithiasis". Biochemistry. 43 (24): 7663–71. doi:10.1021/bi0360758. PMID15196008.
↑Edvardsson VO, Palsson R, Sahota A (1993). Pagon RA, Adam MP, Ardinger HH, Wallace SE, Amemiya A, Bean LJ, Bird TD, Fong CT, Mefford HC, Smith RJ, Stephens K (eds.). "Adenine Phosphoribosyltransferase Deficiency". SourceGeneReviews. PMID22934314.
Further reading
Tischfield JA, Engle SJ, Gupta PK, Bye S, Boyadjiev S, Shao C, O'Neill P, Albertini RJ, Stambrook PJ, Sahota AS (1995). "Germline and Somatic Mutation at the APRT Locus of Mice and Man". Purine and Pyrimidine Metabolism in Man VIII. Advances in Experimental Medicine and Biology. Vol.370. pp.661–4. doi:10.1007/978-1-4615-2584-4_137. ISBN978-1-4613-6105-3. PMID7660991.
Takeuchi H, Kaneko Y, Fujita J, Yoshida O (Apr 1993). "A case of a compound heterozygote for adenine phosphoribosyltransferase deficiency (APRT*J/APRT*Q0) leading to 2,8-dihydroxyadenine urolithiasis: review of the reported cases with 2,8-dihydroxyadenine stones in Japan". The Journal of Urology. 149 (4): 824–6. doi:10.1016/s0022-5347(17)36222-5. PMID8455250.
Ludwig H, Kuzmits R, Pietschmann H, Müller MM (Nov 1979). "Enzymes of the purine interconversion system in chronic lymphatic leukemia: decreased purine nucleoside phosphorylase and adenosine deaminase activity". Blut. 39 (5): 309–15. doi:10.1007/BF01014193. PMID116697. S2CID6283377.
Johnson LA, Gordon RB, Emmerson BT (Apr 1977). "Adenine phosphoribosyltransferase: a simple spectrophotometric assay and the incidence of mutation in the normal population". Biochemical Genetics. 15 (3–4): 265–72. doi:10.1007/BF00484458. PMID869896. S2CID41264715.
Chen J, Sahota A, Stambrook PJ, Tischfield JA (Jul 1991). "Polymerase chain reaction amplification and sequence analysis of human mutant adenine phosphoribosyltransferase genes: the nature and frequency of errors caused by Taq DNA polymerase". Mutation Research. 249 (1): 169–76. Bibcode:1991MRFMM.249..169C. doi:10.1016/0027-5107(91)90143-C. PMID2067530.
Gathof BS, Sahota A, Gresser U, Chen J, Stambrook PJ, Tischfield JA, Zöllner N (Dec 1990). "Identification of a splice mutation at the adenine phosphoribosyltransferase locus in a German family". Klinische Wochenschrift. 69 (24): 1152–5. doi:10.1007/BF01815434. PMID2135300. S2CID11791868.
Chen J, Sahota A, Martin GF, Hakoda M, Kamatani N, Stambrook PJ, Tischfield JA (Jun 1993). "Analysis of germline and in vivo somatic mutations in the human adenine phosphoribosyltransferase gene: mutational hot spots at the intron 4 splice donor site and at codon 87". Mutation Research. 287 (2): 217–25. Bibcode:1993MRFMM.287..217C. doi:10.1016/0027-5107(93)90014-7. PMID7685481.
Sahota A, Chen J, Boyadjiev SA, Gault MH, Tischfield JA (May 1994). "Missense mutation in the adenine phosphoribosyltransferase gene causing 2,8-dihydroxyadenine urolithiasis". Human Molecular Genetics. 3 (5): 817–8. doi:10.1093/hmg/3.5.817. PMID7915931.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.