
Gaviiformes is an order of aquatic birds containing the loons or divers and their closest extinct relatives. Modern gaviiformes are found in many parts of North America and northern Eurasia, though prehistoric species were more widespread.

Hesperornithes is an extinct and highly specialized group of aquatic avialans closely related to the ancestors of modern birds. They inhabited both marine and freshwater habitats in the Northern Hemisphere, and include genera such as Hesperornis, Parahesperornis, Baptornis, Enaliornis, and Potamornis, all strong-swimming, predatory divers. Many of the species most specialized for swimming were completely flightless. The largest known hesperornithean, Canadaga arctica, may have reached a maximum adult length of 2.2 metres (7.2 ft).

Ichthyornis is an extinct genus of toothy seabird-like ornithuran from the late Cretaceous period of North America. Its fossil remains are known from the chalks of Alberta, Alabama, Kansas, New Mexico, Saskatchewan, and Texas, in strata that were laid down in the Western Interior Seaway during the Turonian through Campanian ages, about 95–83.5 million years ago. Ichthyornis is a common component of the Niobrara Formation fauna, and numerous specimens have been found.
Odontornithes is an obsolete and disused taxonomic term proposed by Othniel Charles Marsh for birds possessing teeth, notably the genera Hesperornis and Ichthyornis from the Cretaceous deposits of Kansas.

Gansus is a genus of aquatic birds that lived during the Aptian age of the Early Cretaceous (Aptian-Albian) period in what are now Gansu and Liaoning provinces, western China. The rock layers from which their fossils have been recovered are dated to 120 million years ago. It was first described in 1984 on the basis of an isolated left leg. It is the oldest-known member of the Ornithurae, the group which includes modern birds (Neornithes) and extinct related groups, such as Ichthyornis and Hesperornithes.
Ichthyornithes is an extinct group of toothed avialan dinosaurs very closely related to the common ancestor of all modern birds. They are known from fossil remains found throughout the late Cretaceous period of North America, though only two genera, Ichthyornis and Janavis, are represented by complete enough fossils to have been named. Ichthyornitheans became extinct at the Cretaceous–Paleogene boundary, along with enantiornitheans, all other non-avian dinosaurs, and many other animal and plant groups.

Nyctosaurus is a genus of nyctosaurid pterosaur from the Late Cretaceous period of what is now the Niobrara Formation of the mid-western United States, which, during the time Nyctosaurus was alive, was covered in an extensive shallow sea. Some remains belonging to a possible Nyctosaurus species called "N." lamegoi have been found in Brazil, though it likely belongs to a different genus Simurghia. The genus Nyctosaurus has had numerous species referred to it, though how many of these may actually be valid requires further study. At least one species possessed an extraordinarily large antler-like cranial crest.

Baptornis is a genus of flightless, aquatic birds from the Late Cretaceous, some 87-80 million years ago. The fossils of Baptornis advenus, the type species, were discovered in Kansas, which at its time was mostly covered by the Western Interior Seaway, a shallow shelf sea. It is now known to have also occurred in today's Sweden, where the Turgai Strait joined the ancient North Sea; possibly, it occurred in the entire Holarctic.
Polarornis is a genus of prehistoric bird, possibly an anserimorph. It contains a single species Polarornis gregorii, known from incomplete remains of one individual found on Seymour Island, Antarctica, in rocks which are dated to the Late Cretaceous.

Benjamin Franklin Mudge was an American lawyer, geologist and teacher. Briefly the mayor of Lynn, Massachusetts, he later moved to Kansas where he was appointed the first State Geologist. He led the first geological survey of the state in 1864, and published the first book on the geology of Kansas. He lectured extensively, and was department chair at the Kansas State Agricultural College.
Neogaeornis is a controversial prehistoric genus of diving bird. The single known species, Neogaeornis wetzeli, was described from fossils found in the Campanian to Maastrichtian Quiriquina Formation of Chile. It lived about 70-67 million years ago. It remains known from the single tarsometatarsus described in 1929 by Lambrecht, and today housed in the Paläontologisches Institut und Museum in Kiel, Germany.

Edmontosaurus regalis is a species of comb-crested hadrosaurid dinosaur. Fossils of E. regalis have been found in rocks of western North America that date from the late Campanian age of the Cretaceous Period 73 million years ago, but it may have possibly lived into the early Maastrichtian.
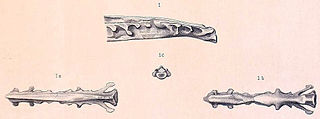
Apatornis is a genus of ornithuran dinosaurs endemic to North America during the late Cretaceous. It currently contains a single species, Apatornis celer, which lived around the Santonian-Campanian boundary, dated to about 83.5 million years ago. The remains of this species were found in the Smoky Hill Chalk of the Niobrara Formation in Kansas, United States. It is known from a single fossil specimen: a synsacrum, the fused series of vertebrae over the hips.

Parahesperornis is a genus of prehistoric flightless birds from the Late Cretaceous. Its range in space and time may have been extensive, but its remains are rather few and far between, at least compared with its contemporary relatives in Hesperornis. Remains are known from central North America, namely the former shallows of the Western Interior Seaway in Kansas. Found only in the upper Niobrara Chalk, these are from around the Coniacian-Santonian boundary, 85–82 million years ago (mya).
Limenavis is a genus of ornithuran dinosaurs from the Late Cretaceous. It lived about 70 million years ago, around the Campanian-Maastrichtian boundary. Known from several broken bones, the remains of the only known species Limenavis patagonica were found in rocks of the "lower member" of the Allen Formation at Salitral Moreno, 20 km south of General Roca, Río Negro (Argentina). It is one of the closest relatives, in the fossil record, of the modern birds.
Ornithurae is a natural group that includes modern birds and their very close relatives such as Ichthyornis and the hesperornithines. This clade is defined in the PhyloCode by Juan Benito and colleagues in 2022 as "the smallest clade containing Ichthyornis dispar, Hesperornis regalis, and Vultur gryphus".
Brodavis is a genus of freshwater hesperornithiform birds known from the Late Cretaceous of North America and Asia. It was first described and named by Larry D. Martin, Evgeny N. Kurochkin and Tim T. Tokaryk in 2012 and assigned to a new monogeneric family, Brodavidae. Four species were described and assigned to Brodavis.

Tingmiatornis is a genus of flighted and possibly diving ornithurine dinosaur from the High Arctic of Canada. The genus contains a single species, T. arctica, described in 2016, which lived during the Turonian epoch of the Cretaceous.
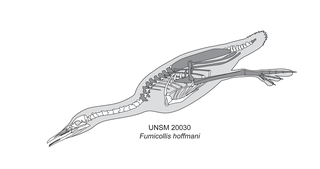
Fumicollis is a genus of prehistoric flightless birds from the Late Cretaceous (Coniacian-Santonian) Niobrara Chalk of Kansas.

Jormungandr is an extinct genus of mosasaurid squamates from the early Campanian Pierre Shale of North Dakota, United States. The genus contains a single species, J. walhallaensis, known from a nearly-complete skull and partial skeleton. Jormungandr was a medium-sized mosasaur, at around 6–8 metres (20–26 ft) long, and its skeletal anatomy exhibits a mix of features seen in both basal and derived mosasaurines.


















{kind=link}
{kind=link}
{kind=link}