| Name | Year | Formation | Location | Notes | Images |
|---|
| Abditosaurus | 2022 | Tremp Formation (Late Cretaceous, Maastrichtian) |  Spain Spain | Larger and distantly related to other European sauropods, suggesting it was an immigrant from another continent [1] |  |
| Acanthopholis | 1867 | Chalk Group (Late Cretaceous, Cenomanian) |  United Kingdom United Kingdom
( England) England) | Possessed keeled oval scutes as well as long spines |  |
| Adynomosaurus | 2019 | Tremp Formation (Late Cretaceous, Maastrichtian) | Spain | Relatively basal despite its late age [2] |  |
| Aepisaurus | 1852 | Grès verts hevétiques (Early Cretaceous, Albian) |  France France | Known only from a humerus which shares some features with camarasaurids and titanosaurs [3] |  |
| Ajkaceratops | 2010 | Csehbánya Formation (Late Cretaceous, Santonian) |  Hungary Hungary | Known for its sharply hooked upper beak |  |
| Allosaurus | 1877 | Lourinhã Formation (Late Jurassic, Kimmeridgian) |  Portugal Portugal | Three species, including the type, are known from the United States. The European species may be a synonym of A. fragilis, [4] [5] [6] but at least one study suggests that it is indeed valid [7] |  |
| Alocodon | 1973 | Cabaços Formation (Late Jurassic, Oxfordian) | Portugal | Had vertical grooves on its teeth |  |
| Altispinax | 1923 | Wadhurst Clay Formation (Early Cretaceous, Valanginian) | United Kingdom
( England) | Possessed elongated neural spines that may have supported a hump-like structure as in Concavenator |  |
| Amanzia | 2020 | Reuchenette Formation (Late Jurassic, Kimmeridgian) |  Switzerland Switzerland | One bone preserves fossilized cartilage. [8] Originally believed to be a species of Ornithopsis or Cetiosauriscus |  |
| Ampelosaurus | 1995 | Argiles et Grès à Reptiles Formation, Grès de Labarre, Gres de Saint-Chinian, Marnes Rouges Inférieures Formation (Late Cretaceous, Maastrichtian) | France | Had three different types of osteoderms, including spines, plates and bulbs |  |
| Anoplosaurus | 1879 | Cambridge Greensand (Early Cretaceous, Albian) | United Kingdom
( England) | Considered an ankylosaur despite the fact no osteoderms have been found. Historically suggested to be an ornithopod | 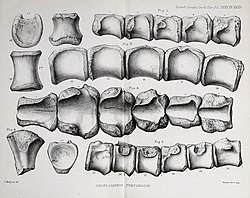 |
| Aragosaurus | 1987 | Villar del Arzobispo Formation (Early Cretaceous, Berriasian) | Spain | Reportedly had a unique ischium |  |
| Arcovenator | 2014 | Argiles et Grès à Reptiles Formation (Late Cretaceous, Campanian) | France | Most closely related to abelisaurids from India and Madagascar [9] |  |
| Arenysaurus | 2009 | Tremp Formation (Late Cretaceous, Maastrichtian) | Spain | One of the last lambeosaurines prior to their extinction |  |
| Aristosuchus | 1887 | Wealden Group (Early Cretaceous, Barremian) | United Kingdom
( England) | May have had long claws based on referred unguals |  |
| Asylosaurus | 2007 | Avon Fissure Fill (Late Triassic, Rhaetian) | United Kingdom
( England) | Remains originally identified as Thecodontosaurus |  |
| Atsinganosaurus | 2010 | Argiles et Grès à Reptiles Formation (Late Cretaceous, Maastrichtian) | France | Belongs to a unique clade of southwestern European titanosaurs [10] |  |
| Aviatyrannis | 2003 | Alcobaça Formation (Late Jurassic, Oxfordian to Tithonian) | Portugal | Although originally described as a tyrannosauroid, it may in fact be the oldest known ornithomimosaur [11] | 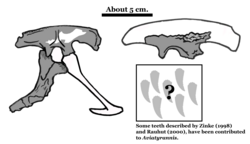 |
| Barilium | 2010 | Wadhurst Clay Formation (Early Cretaceous, Valanginian) | United Kingdom
( England) | Robust with strong vertebrae and short neural spines |  |
| Baryonyx | 1986 | Weald Clay (Early Cretaceous, Barremian) | United Kingdom
( England) | One specimen was found with the remains of fish and a juvenile iguanodont in its stomach, suggesting it was a generalist predator [12] |  |
| Betasuchus | 1932 | Maastricht Formation (Late Cretaceous, Maastrichtian) |  Netherlands Netherlands | The first terrestrial vertebrate named from the Maastrichtian stage [13] |  |
| Blasisaurus | 2010 | Arén Formation (Late Cretaceous, Maastrichtian) | Spain | Only known from a partial skull but can be distinguished from contemporary lambeosaurines [14] |  |
| Bothriospondylus | 1875 | Kimmeridge Clay (Late Jurassic, Kimmeridgian) | United Kingdom
( England) | Several specimens have been assigned to this genus, but most of them have been reclassified into different genera |  |
| Bradycneme | 1975 | Sânpetru Formation (Late Cretaceous, Maastrichtian) |  Romania Romania | Potentially an alvarezsaurid [8] |  |
| Brighstoneus | 2021 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Possessed a long snout tipped with a low bump |  |
| Burianosaurus | 2017 | Peruc-Korycany Formation (Late Cretaceous, Cenomanian) |  Czech Republic Czech Republic | May be closely allied to the rhabdodontids [15] |  |
| Calamosaurus | 1891 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Only known from two cervical vertebrae. Several bones have been referred to it over the years but there is no way to prove all of them belonged to the same genus | 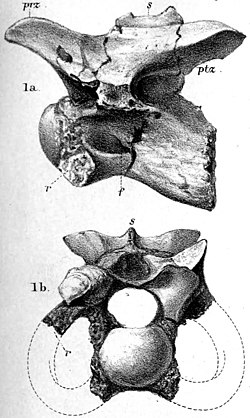 |
| Calamospondylus | 1866 | Wessex Formation (Early Cretaceous, Hauterivian) | United Kingdom
( England) | Only definitively known from a sacrum and associated pelvic elements |  |
| Caletodraco | 2024 | Chalk of the Pays de Caux (Late Cretaceous, Cenomanian) | France | Possibly the first known non-South American furileusaurian abelisaurid [16] |  |
| Callovosaurus | 1980 | Oxford Clay (Middle Jurassic, Callovian) | United Kingdom
( England) | The oldest known dryosaurid [17] |  |
| Calvarius | 2023 | Tremp Formation (Late Cretaceous, Maastrichtian) | Spain | May have been a cursorial biped similar to basal ornithopods despite being more derived |  |
| Camarillasaurus | 2014 | Camarillas Formation (Early Cretaceous, Barremian) | Spain | Described as a ceratosaur [18] but has since been reinterpreted as a spinosaurid [19] [20] |  |
| Camelotia | 1985 | Westbury Formation (Late Triassic to Early Jurassic, Rhaetian to Hettangian) | United Kingdom
( England) | One of the largest known non-sauropod sauropodomorphs |  |
| Canardia | 2013 | Marnes d'Auzas Formation (Late Cretaceous, Maastrichtian) | France | May have been a close relative of Aralosaurus |  |
| Cardiodon | 1841 | Forest Marble Formation (Middle Jurassic, Bathonian) | United Kingdom
( England) | The first sauropod ever named. [21] Known only from a tooth | 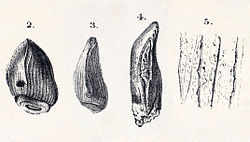 |
| Cariocecus | 2025 | Papo Seco Formation (Early Cretaceous, Barremian) | Portugal | Represents the first Portuguese iguanodontian skull [22] |  |
| Ceratosuchops | 2021 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Differs from Baryonyx in subtle details of its skull |  |
| Cetiosauriscus | 1927 | Oxford Clay (Middle Jurassic, Callovian) | United Kingdom
( England) | Has been assigned to a variety of positions around Eusauropoda [23] [24] [25] |  |
| Cetiosaurus | 1841 | Rutland Formation (Middle Jurassic, Bajocian to Bathonian) | United Kingdom
( England) | The first sauropod known from decent remains. Once believed to be a large seagoing animal |  |
| Chondrosteosaurus | 1876 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | The air sacs in its vertebrae were originally believed to be filled with cartilage | 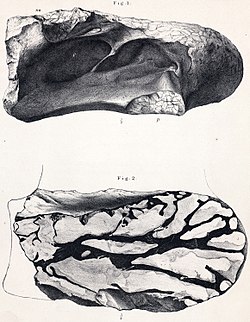 |
| Compsognathus | 1859 | Alcobaça Formation?, Canjuers Lagerstätte, Solnhofen Limestone (Late Jurassic, Tithonian) | France
 Germany Germany
Portugal? | One of the smallest known non-avian dinosaurs |  |
| Comptonatus | 2024 | Wessex Formation (Early Cretaceous, Barremian to Aptian) | United Kingdom
( England) | One of the most completely known ornithopod taxa from the Isle of Wight |  |
| Concavenator | 2010 | La Huérguina Formation (Early Cretaceous, Barremian) | Spain | Preserves bumps on its ulna which have been interpreted as quill knobs, [26] although they might have been muscle attachments instead [27] |  |
| Craspedodon | 1883 | Lonzée Member (Late Cretaceous, Santonian) |  Belgium Belgium | May be the first neoceratopsian known from Europe [28] |  |
| Craterosaurus | 1874 | Woburn Sands Formation (Early Cretaceous, Aptian to Albian) | United Kingdom
( England) | Potentially synonymous with Regnosaurus |  |
| Cruxicheiros | 2010 | Chipping Norton Limestone (Middle Jurassic, Bathonian) | United Kingdom
( England) | Inconsistent in phylogenetic placement |  |
| Cryptosaurus | 1869 | Ampthill Clay (Late Jurassic, Oxfordian) | United Kingdom
( England) | Only known from a single femur |  |
| Cumnoria | 1888 | Kimmeridge Clay (Late Jurassic, Kimmeridgian) | United Kingdom
( England) | May be a species of Camptosaurus [29] or something a little more basal [30] |  |
| Dacentrurus | 1902 | Alcobaça Formation, Argiles d'Octeville, Kimmeridge Clay, Lourinhã Formation, Villar del Arzobispo Formation (Late Jurassic to Early Cretaceous, Kimmeridgian to Berriasian) | France
Portugal
Spain
United Kingdom
( England) | Known from abundant remains. Lived in a broad range |  |
| Demandasaurus | 2011 | Castrillo de la Reina Formation (Early Cretaceous, Barremian to Aptian) | Spain | Most closely related to African rebbachisaurids, suggesting a faunal exchange [31] |  |
| Dinodocus | 1884 | Lower Greensand Group (Early Cretaceous, Barremian) | United Kingdom
( England) | The only known humerus is almost complete, missing only small portions |  |
| Dolichosuchus | 1932 | Löwenstein Formation (Late Triassic, Norian) | Germany | Originally classified as a stem-crocodile |  |
| Dornraptor | 2024 | Blue Lias?/Charmouth Mudstone Formation? (Early Jurassic, Sinemurian) | United Kingdom
( England) | Originally thought to belong to Scelidosaurus , [32] its remains were then informally named "Merosaurus" before their formal description in 2024 [33] |  |
| Draconyx | 2001 | Lourinhã Formation (Late Jurassic, Tithonian) | Portugal | May have been a member of an ornithopod clade that did not appear in North America, unlike other groups of Late Jurassic animals [34] |  |
| Dracopelta | 1980 | Lourinhã Formation (Late Jurassic, Tithonian) | Portugal | The structure of its limbs suggests it might have had a cursorial lifestyle [35] | |
| Dracoraptor | 2016 | Blue Lias (Early Jurassic, Hettangian) | United Kingdom
( Wales) Wales) | May have been a shore dweller due to its island habitat [36] |  |
| Dromaeosauroides | 2003 | Jydegaard Formation (Early Cretaceous, Berriasian) |  Denmark Denmark | A referred coprolite has been found which contains fish remains, but it could instead belong to a turtle [37] |  |
| Dubreuillosaurus | 2005 | Calcaire de Caen (Middle Jurassic, Bathonian) | France | Would have lived in a coastal mangrove swamp |  |
| Duriatitan | 2010 | Kimmeridge Clay (Late Jurassic, Kimmeridgian) | United Kingdom
( England) | Originally named as a species of Cetiosaurus |  |
| Duriavenator | 2008 | Inferior Oolite (Middle Jurassic, Bajocian) | United Kingdom
( England) | The basalmost known member of the Megalosaurinae, which aligns with its stratigraphic position [38] |  |
| Echinodon | 1861 | Purbeck Group (Early Cretaceous, Berriasian) | United Kingdom
( England) | Originally misidentified as a herbivorous lizard |  |
| Efraasia | 1973 | Löwenstein Formation (Late Triassic, Norian) | Germany | Some remains assigned to this genus were originally classified in separate genera |  |
| Emausaurus | 1990 | Ciechocinek Formation (Early Jurassic, Toarcian) | Germany | One of the oldest and basalmost thyreophorans |  |
| Eotyrannus | 2001 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Possessed grasping hands with three long fingers |  |
| Eousdryosaurus | 2014 | Lourinhã Formation (Late Jurassic, Kimmeridgian) | Portugal | Described as a dryosaurid [39] but one study suggests a close relationship with elasmarians [40] | |
| Erectopus | 1923 | La Penthiève Beds (Early Cretaceous, Albian) | France | One of the youngest known European carnosaurs | 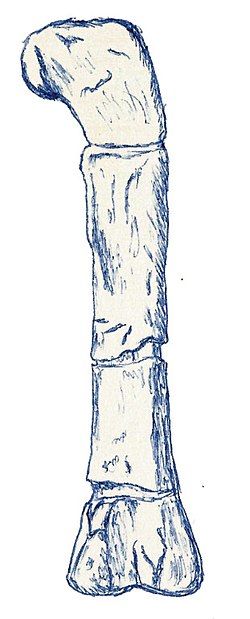 |
| Eucamerotus | 1872 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Preserves extensive evidence of pneumatization | |
| Eucercosaurus | 1879 | Cambridge Greensand (Early Cretaceous, Albian) | United Kingdom
( England) | Sometimes considered an ankylosaur but one study assigns it to Iguanodontia [41] |  |
| Euronychodon | 1991 | Argiles et sables de Taveiro (Late Cretaceous, Campanian to Maastrichtian) | Portugal | Only known from teeth. Another species has been found in Uzbekistan |  |
| Europasaurus | 2006 | Süntel Formation (Late Jurassic, Kimmeridgian) | Germany | Much smaller than other sauropods due to its isolated island habitat [42] |  |
| Europatitan | 2017 | Castrillo de la Reina Formation (Early Cretaceous, Barremian to Aptian) | Spain | Some of this genus' remains include several vertebrae. The specific name, eastwoodi, honors director Clint Eastwood |  |
| Europelta | 2013 | Escucha Formation (Early Cretaceous, Albian) | Spain | Almost the entire skeleton is known |  |
| Eustreptospondylus | 1964 | Oxford Clay (Middle Jurassic to Late Jurassic, Callovian to Oxfordian) | United Kingdom
( England) | May have swum between islands similar to a Komodo dragon [43] |  |
| Ferenceratops | 2026 | Densuș-Ciula Formation, Sânpetru Formation (Late Cretaceous, Maastrichtian) | Romania | Originally named as a species of Zalmoxes but it was reclassified as a ceratopsian |  |
| Foskeia | 2026 | Castrillo de la Reina Formation (Early Cretaceous, Barremian to Aptian) | Spain | Before its formal description, it had been nicknamed the "Vegagete ornithopod". One of the smallest known ornithopods |  |
| Fylax | 2021 | Figuerola Formation (Late Cretaceous, Maastrichtian) | Spain | Lived very late despite its comparatively basal position [44] |  |
| Galvesaurus | 2005 | Villar del Arzobispo Formation (Late Jurassic, Tithonian) | Spain | Also spelled Galveosaurus. Two sets of paleontologists named the same fossil seemingly unaware of each other's work, although there is evidence that one of them had plaigiarized the others, but misspelled the name [45] | |
| Garrigatitan | 2021 | Argiles et Grès à Reptiles Formation (Late Cretaceous, Campanian) | France | Known from remains of both adults and subadults | |
| Garumbatitan | 2023 | Arcillas de Morella Formation (Early Cretaceous, Barremian) | Spain | Had a reduced claw on the third toe |  |
| Genusaurus | 1995 | Bevons Beds (Early Cretaceous, Albian) | France | Has been suggested to be either a noasaurid [46] or an abelisaurid [47] | |
| Gideonmantellia | 2012 | Camarillas Formation (Early Cretaceous, Barremian) | Spain | Originally misidentified as a specimen of Hypsilophodon | 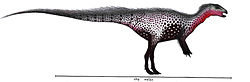 |
| Gigantosaurus | 1869 | Kimmeridge Clay (Late Jurassic, Kimmeridgian) | United Kingdom
( England) | May have possessed osteoderms |  |
| Haestasaurus | 2015 | Wealden Group (Early Cretaceous, Berriasian to Valanginian) | United Kingdom
( England) | Preserves impressions of differently-sized hexagonal scales |  |
| Halticosaurus | 1908 | Löwenstein Formation (Late Triassic, Norian) | Germany | Historically conflated with the bones of unrelated animals | |
| Heptasteornis | 1975 | Sânpetru Formation (Late Cretaceous, Maastrichtian) | Romania | Once believed to be a giant prehistoric owl |  |
| Hesperonyx | 2024 | Lourinhã Formation (Late Jurassic, Kimmeridgian) | Portugal | Only known from the remains of the fore- and hindlimbs |  |
| Histriasaurus | 1998 | Unnamed formation (Early Cretaceous, Hauterivian to Barremian) |  Croatia Croatia | Despite being discovered in Europe, it may have lived between southern Europe and Africa in life [48] |  |
| Horshamosaurus | 2015 | Weald Clay (Early Cretaceous, Barremian) | United Kingdom
( England) | A supposed tibia has been reinterpreted as an ischium [49] |  |
| Hungarosaurus | 2005 | Csehbánya Formation (Late Cretaceous, Santonian) | Hungary | Possessed an elevated shoulder which may be an adaptation to high-browsing [50] |  |
| Hylaeosaurus | 1833 | Bückeberg Formation?, Grinstead Clay Formation, Tunbridge Wells Sand Formation (Early Cretaceous, Valanginian) | Germany?
United Kingdom
( England) | One of the three animals originally used to define the Dinosauria, along with Iguanodon and Megalosaurus |  |
| Hypselosaurus | 1869 | Argiles et Grès à Reptiles Formation (Late Cretaceous, Maastrichtian) | France | Several spherical eggs have been attributed to this taxon |  |
| Hypselospinus | 2010 | Wadhurst Clay Formation (Early Cretaceous, Valanginian) | United Kingdom
( England) | Had elongated neural spines projecting from the top of its vertebrae |  |
| Hypsilophodon | 1869 | Wessex Formation (Early Cretaceous, Hauterivian to Barremian) | United Kingdom
England | May have been a deer-like low browser that fed on young shoots and roots |  |
| Iberospinus | 2022 | Papo Seco Formation (Early Cretaceous, Barremian) | Portugal | Basal yet already displays some adaptations for a semiaquatic lifestyle |  |
| Iguanodon | 1825 | Arcillas de Morella Formation, Camarillas Formation, Sainte-Barbe Clays Formation, Wadhurst Clay Formation, Weald Clay, Wealden Group, Wessex Formation (Early Cretaceous, Barremian to Aptian) | Belgium
Germany?
Spain
United Kingdom
( England) | Multiple remains are known which make it one of the best known dinosaurs | 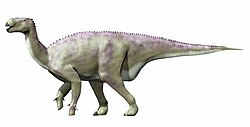 |
| Iliosuchus | 1932 | Taynton Limestone Formation (Middle Jurassic, Bathonian) | United Kingdom
( England) | Only known from three ilia |  |
| Istiorachis | 2025 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Possessed a sail-like structure formed by elongated neural spines |  |
| Iuticosaurus | 1993 | Upper Greensand Formation, Wessex Formation (Early Cretaceous to Late Cretaceous, Berriasian to Cenomanian) | United Kingdom
( England) | Two species have been named, each from a single caudal vertebra |  |
| Juratyrant | 2013 | Kimmeridge Clay (Late Jurassic, Tithonian) | United Kingdom
( England) | Originally named as a species of Stokesosaurus |  |
| Juravenator | 2006 | Torleite Formation (Late Jurassic, Kimmeridgian) | Germany | Impressions of both scales and feathers are known. The tail preserves structures that may be integumentary sense organs like those of crocodiles, which it may have used to prey on fish at night [51] |  |
| Kryptohadros | 2026 | Densuș-Ciula Formation (Late Cretaceous, Maastrichtian) | Romania | Represents the family Telmatosauridae, a clade of exclusively southeastern European hadrosauroids |  |
| Lexovisaurus | 1957 | Oxford Clay (Middle Jurassic, Callovian) | United Kingdom
( England) | Its pelvis was greatly enlarged |  |
| Liliensternus | 1984 | Trossingen Formation (Late Triassic, Norian to Rhaetian) | Germany | Although commonly depicted with a head crest, there is no evidence for such a feature |  |
| Lirainosaurus | 1999 | Marnes Rouges Inférieures Formation?, Sierra Perenchiza Formation, Sobrepena Formation (Late Cretaceous, Campanian to Maastrichtian) | France?
Spain | For a titanosaur, it was small and had a relatively gracile build |  |
| Lohuecotitan | 2016 | Villalba de la Sierra Formation (Late Cretaceous, Campanian to Maastrichtian) | Spain | May have had "bulb-and-root"-type osteoderms, which are abundant at the type locality [52] |  |
| Lophostropheus | 2007 | Moon-Airel Formation (Late Triassic to Early Jurassic, Rhaetian to Hettangian) | France | The only substantially well-known theropod from the Triassic-Jurassic boundary [53] | |
| Loricatosaurus | 2008 | Marnes à Belemnopsis latesulcatus Formation, Oxford Clay (Middle Jurassic, Callovian) | France
United Kingdom
( England) | Had narrow, flat plates on its back and round, pointed spines that ran along the tail |  |
| Losillasaurus | 2001 | Villar del Arzobispo Formation (Late Jurassic to Early Cretaceous?, Kimmeridgian to Berriasian?) | Spain | Was heterodont, having four types of teeth, one of which was heart-shaped [54] |  |
| Lourinhanosaurus | 1998 | Lourinhã Formation (Late Jurassic, Kimmeridgian to Tithonian) | Portugal | More than one hundred eggs have been referred to this taxon |  |
| Lourinhasaurus | 1998 | Lourinhã Formation (Late Jurassic, Kimmeridgian) | Portugal | Closely related to Camarasaurus but with proportionately longer forelimbs |  |
| Lusotitan | 2003 | Lourinhã Formation (Late Jurassic, Tithonian) | Portugal | Originally named as a European species of Brachiosaurus |  |
| Lusovenator | 2020 | Lourinhã Formation (Late Jurassic to Early Cretaceous, Kimmeridgian to Berriasian) | Portugal | The oldest carcharodontosaurian known from Eurasia |  |
| Macrurosaurus | 1869 | Cambridge Greensand, Chalk Group? (Early Cretaceous, Albian) | United Kingdom
( England) | Only known from a series of caudal vertebrae |  |
| Magnamanus | 2016 | Golmayo Formation (Early Cretaceous, Hauterivian to Barremian) | Spain | Possessed relatively enlarged hands | |
| Magnosaurus | 1932 | Inferior Oolite (Middle Jurassic, Bajocian) | United Kingdom
( England) | Confusingly, a referred specimen was simultaneously named as a species of this genus and of Sarcosaurus |  |
| Magyarosaurus | 1932 | Sânpetru Formation (Late Cretaceous, Maastrichtian) | Romania | An insular dwarf titanosaur that was one of the smallest of its group |  |
| Mantellisaurus | 2007 | Arcillas de Morella Formation, La Huérguina Formation, Lower Greensand Group, Sainte-Barbe Clays Formation, Vectis Formation, Wessex Formation (Early Cretaceous, Barremian to Aptian) | Belgium
Germany?
Spain
United Kingdom
( England) | Several specimens are known. Distinguishable from the contemporary Iguanodon by its more gracile build |  |
| Marmarospondylus | 1875 | Forest Marble Formation (Middle Jurassic, Bathonian) | United Kingdom
( England) | Usually assigned to the genus Bothriospondylus , but this cannot be confirmed |  |
| Matheronodon | 2017 | Argiles et Grès à Reptiles Formation (Late Cretaceous, Campanian) | France | Had extremely specialized dentition that may have been an adaptation to feeding on tough monocot plants [55] |  |
| Megalosaurus | 1824 | Chipping Norton Limestone, Taynton Limestone Formation (Middle Jurassic, Bathonian) | United Kingdom
( England) | The first non-avian dinosaur scientifically named and described |  |
| Metriacanthosaurus | 1964 | Oxford Clay (Late Jurassic, Oxfordian) | United Kingdom
( England) | Possessed relatively tall neural spines for a carnosaur |  |
| Miragaia | 2009 | Lourinhã Formation (Late Jurassic, Kimmeridgian to Tithonian) | Portugal | Had an extremely elongated neck made up of seventeen vertebrae. May be synonymous with Dacentrurus [56] [57] |  |
| Mochlodon | 1881 | Csehbánya Formation, Gosau Group (Late Cretaceous, Santonian to Campanian) |  Austria Austria
Hungary | Originally named as a species of Iguanodon |  |
| Morelladon | 2015 | Arcillas de Morella Formation (Early Cretaceous, Barremian) | Spain | Possessed a neural spine sail lower than that of Ouranosaurus |  |
| Morinosaurus | 1874 | Unnamed formation (Late Jurassic, Kimmeridgian) | France | Poorly known |  |
| Neosodon | 1885 | Sables et Grès à Trigonia gibbosa (Late Jurassic, Tithonian) | France | No species are assigned to this genus. Popularly associated with "Iguanodon" praecursor but is in fact a separate taxon [58] |  |
| Neovenator | 1996 | Wessex Formation (Early Cretaceous, Hauterivian to Barremian) | United Kingdom
( England) | Had a complex series of neurovascular canals lining its snout [59] |  |
| Newtonsaurus | 2025 | Lilstock Formation (Late Triassic, Rhaetian) | United Kingdom
( Wales) | Once considered indeterminate within Archosauria |  |
| Normanniasaurus | 2013 | Poudingue Ferrugineux (Early Cretaceous, Albian) | France | Represents a European radiation of basal titanosaurs [60] | |
| Notatesseraeraptor | 2019 | Klettgau Formation (Late Triassic, Norian) | Switzerland | Combines features of different groups of basal theropods |  |
| Nuthetes | 1854 | Lulworth Formation (Early Cretaceous, Berriasian) | United Kingdom
( England) | Only known from jaws and teeth. Traditionally identified as a dromaeosaurid but it might be a proceratosaurid instead [61] |  |
| Obelignathus | 2025 | Argiles et Grès à Reptiles Formation (Late Cretaceous, Campanian to Maastrichtian) | France | Originally named as a species of Rhabdodon , but was given its own genus due to morphological and phylogenetic differences [62] |  |
| Oblitosaurus | 2023 | Villar del Arzobispo Formation (Late Jurassic, Kimmeridgian to Tithonian) | Spain | The largest ornithopod known from the Late Jurassic of Europe |  |
| Oceanotitan | 2019 | Lourinhã Formation (Late Jurassic, Kimmeridgian) | Portugal | Potentially the oldest known somphospondylian [63] |  |
| Ohmdenosaurus | 1978 | Posidonia Shale (Early Jurassic, Toarcian) | Germany | Originally misidentified as a plesiosaur |  |
| Oplosaurus | 1852 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | The holotype tooth was pointed, which led to its misidentification as a carnivorous reptile [64] | 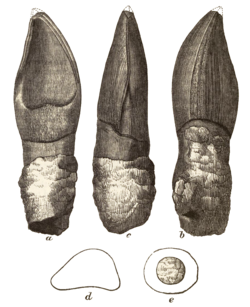 |
| Ornithodesmus | 1887 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Historically conflated with the remains of the pterosaur Istiodactylus |  |
| Ornithopsis | 1870 | Wealden Group (Early Cretaceous, Barremian) | Germany?
United Kingdom
( England) | Originally believed to be an intermediate form between birds, pterosaurs and dinosaurs |  |
| Orthomerus | 1883 | Maastricht Formation (Late Cretaceous, Maastrichtian) | Netherlands | Potentially dubious and undiagnostic [65] |  |
| Ostromia | 2017 | Painten Formation (Late Jurassic, Tithonian) | Germany | Considered a small pterosaur until it was redescribed as a specimen of Archaeopteryx in 1970. [66] Some of its features are similar to those of Anchiornis |  |
| Owenodon | 2009 | Lulworth Formation (Early Cretaceous, Berriasian) | United Kingdom
( England) | Has been assigned to Iguanodon and Camptosaurus before it received its own genus | 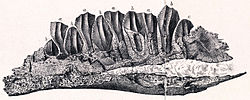 |
| Paludititan | 2010 | Sânpetru Formation (Late Cretaceous, Maastrichtian) | Romania | Some of its bones are identical to those of Magyarosaurus , but their synonymy cannot be confirmed |  |
| Pararhabdodon | 1993 | Tremp Formation (Late Cretaceous, Maastrichtian) | Spain | The first lambeosaurine identified from Europe |  |
| Pareisactus | 2019 | Tremp Formation (Late Cretaceous, Maastrichtian) | Spain | Represented by a single scapula recovered from a hadrosaur bonebed | 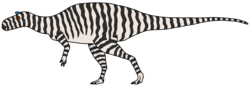 |
| Paulodon | 2025 | Camarillas Formation (Early Cretaceous, Barremian) | Spain | Previously assigned to the contemporary Iguanodon |  |
| Pelecanimimus | 1994 | La Huérguina Formation (Early Cretaceous, Barremian) | Spain | Preserves extensive soft tissue impressions revealing the presence of a keratinous head crest and a pelican-like gular pouch |  |
| Pelorosaurus | 1850 | Tunbridge Wells Sand Formation (Early Cretaceous, Hauterivian) | United Kingdom
( England) | The first sauropod identified as a terrestrial animal |  |
| Pendraig | 2021 | Pant-y-Ffynnon Quarry (Late Triassic, Rhaetian) | United Kingdom
( Wales) | Would have lived on a dry limestone island [67] |  |
| Petrustitan | 2025 | Sânpetru Formation (Late Cretaceous, Maastrichtian) | Romania | Formerly identified as a species of Magyarosaurus , but it is markedly different from that genus |  |
| Phyllodon | 1973 | Alcobaça Formation (Late Jurassic, Kimmeridgian) | Portugal | The front and back of its teeth were asymmetrical |  |
| Piveteausaurus | 1977 | Marnes de Dives (Middle Jurassic, Callovian) | France | Has been suggested to be a species of Proceratosaurus |  |
| Plateosaurus | 1837 | Klettgau Formation, Löwenstein Formation, Lunde Formation, Trossingen Formation (Late Triassic, Norian) | France?
Germany
 Norway Norway
Switzerland | Known from over a hundred skeletons, several of them nearly complete |  |
| Pneumatoraptor | 2010 | Csehbánya Formation (Late Cretaceous, Santonian) | Hungary | One study suggests a position as a possible early palaeognath [68] |  |
| Poekilopleuron | 1836 | Calcaire de Caen (Middle Jurassic, Bathonian) | France | Its holotype was found alongside fish remains [69] |  |
| Polacanthus | 1865 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Possessed a flat sacral shield dotted with small bumps |  |
| Portellsaurus | 2021 | Margas de Mirambell Formation (Early Cretaceous, Barremian) | Spain | Closely related to Ouranosaurus [70] |  |
| Priodontognathus | 1875 | Lower Calcareous Grit (Late Jurassic, Oxfordian) | United Kingdom
( England) | Only known from a single maxilla with teeth | |
| Proa | 2012 | Escucha Formation (Early Cretaceous, Albian) | Spain | The tip of its jaw was shaped like the bow of a ship |  |
| Proceratosaurus | 1926 | Great Oolite Group (Middle Jurassic, Bathonian) | United Kingdom
( England) | Preserves a small horn on its snout which may have anchored a crest as in the related Guanlong |  |
| Procompsognathus | 1913 | Löwenstein Formation (Late Triassic, Norian) | Germany | Has been suggested to be non-dinosaurian |  |
| Protathlitis | 2023 | Arcillas de Morella Formation (Early Cretaceous, Barremian) | Spain | Described as a basal baryonychine spinosaurid, but a recent study suggested the type material may be chimeric, regarding this genus as probably dubious [20] |  |
| Pterospondylus | 1913 | Trossingen Formation (Late Triassic, Norian) | Germany | Known from only a single, large vertebra |  |
| Pyroraptor | 2000 | Argiles et Grès à Reptiles Formation (Late Cretaceous, Campanian to Maastrichtian) | France | Its holotype specimen was exposed by a forest fire |  |
| Qunkasaura | 2024 | Villalba de la Sierra Formation (Late Cretaceous, Campanian to Maastrichtian) | Spain | May have been closely related to Abditosaurus [71] |  |
| Regnosaurus | 1848 | Tunbridge Wells Sand Formation (Early Cretaceous, Berriasian to Valanginian) | United Kingdom
( England) | Potentially a stegosaur similar to Huayangosaurus [72] | 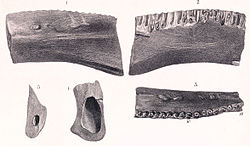 |
| Rhabdodon | 1869 | Grès de Labarre, Gres de Saint-Chinian, Marnes Rouges Inférieures Formation, Villalba de la Sierra Formation (Late Cretaceous, Maastrichtian) | France
Spain | Although most rhabdodontids are insular dwarfs, this genus may represent an instance of island gigantism as it is much larger than other members of its family [73] |  |
| Riabininohadros | 2020 | Unnamed formation (Late Cretaceous, Maastrichtian) |  Ukraine Ukraine | Possessed a femur so unique it has no morphological equivalents across all of Iguanodontia | |
| Riojavenatrix | 2024 | Enciso Group (Early Cretaceous, Barremian to Aptian) | Spain | Originally identified as a specimen of Baryonyx |  |
| Riparovenator | 2021 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Had elongated spines projecting from its caudal vertebrae somewhat similar to those of Spinosaurus |  |
| Ruehleia | 2001 | Trossingen Formation (Late Triassic, Norian) | Germany | Known from a single, nearly complete skeleton |  |
| Saltriovenator | 2018 | Saltrio Formation (Early Jurassic, Sinemurian) |  Italy Italy | The biggest theropod from the Early Jurassic yet known |  |
| Sarcolestes | 1893 | Oxford Clay (Middle Jurassic, Callovian) | United Kingdom
( England) | Originally misidentified as a carnivorous dinosaur |  |
| Sarcosaurus | 1921 | Scunthorpe Mudstone (Early Jurassic, Hettangian to Sinemurian) | United Kingdom
( England) | Has been variously suggested to be in different positions at the base of Neotheropoda [74] [75] [76] [77] |  |
| Scelidosaurus | 1859 | Charmouth Mudstone Formation (Early Jurassic, Sinemurian to Pliensbachian) | United Kingdom
( England) | Carried hundreds of small osteoderms in several rows along its back |  |
| Schleitheimia | 2020 | Klettgau Formation (Late Triassic, Norian) | Switzerland | Possessed a relatively enlarged ilium | |
| Scipionyx | 1998 | Pietraroja Plattenkalk (Early Cretaceous, Albian) | Italy | So well preserved that several internal organs and their positions in life could be accurately reconstructed |  |
| Sciurumimus | 2012 | Torleite Formation (Late Jurassic, Kimmeridgian) | Germany | Had a bushy tail similar to that of a squirrel |  |
| Soriatitan | 2017 | Golmayo Formation (Early Cretaceous, Hauterivian to Barremian) | Spain | The first confirmed brachiosaurid known from Early Cretaceous Europe |  |
| Stenopelix | 1857 | Obernkirchen Sandstein (Early Cretaceous, Berriasian) | Germany | Possibly closely related to basal ceratopsians from Late Jurassic China [78] | 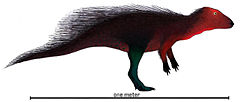 |
| Streptospondylus | 1832 | Marnes de Dives?/Marnes de Villers? (Middle Jurassic to Late Jurassic, Callovian to Oxfordian) | France | Originally believed to represent a marine crocodile |  |
| Struthiosaurus | 1871 | Argiles et Grès à Reptiles Formation, Gosau Group, Sânpetru Formation (Late Cretaceous, Santonian to Maastrichtian) | Austria
France
Hungary?
Romania | Analysis of its braincase suggests poor hearing and a sluggish, solitary lifestyle [79] |  |
| Syngonosaurus | 1879 | Cambridge Greensand (Early Cretaceous, Albian) | United Kingdom
( England) | Usually considered a synonym of Acanthopholis but it has been reinterpreted as an iguanodont [41] |  |
| Tamarro | 2021 | Tremp Formation (Late Cretaceous, Maastrichtian) | Spain | May have been closely related to Asian troodontids [80] |  |
| Tanystrosuchus | 1963 | Löwenstein Formation (Late Triassic, Norian) | Germany | The only known vertebra was once misidentified as a phytosaur | |
| Tarascosaurus | 1991 | Fuvelian Beds (Late Cretaceous, Campanian) | France | If an abelisaurid it would be one of the few northern members of the group |  |
| Tastavinsaurus | 2008 | Forcall Formation, Xert Formation (Early Cretaceous, Barremian) | Spain | Could represent an obscure group of macronarians called Laurasiformes [81] |  |
| Teinurosaurus | 1928 | Mont-Lambert Formation (Late Jurassic, Tithonian) | France | Poorly known |  |
| Telmatosaurus | 1903 | Sânpetru Formation (Late Cretaceous, Maastrichtian) | Romania | One specimen preserves a facial deformity caused by an ameloblastoma [82] |  |
| Tethyshadros | 2009 | Calcare di Aurisina (Late Cretaceous, Campanian) | Italy | Had limbs adapted for high speed but were too short for running [50] |  |
| Thecocoelurus | 1923 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Has been assigned to a variety of theropod groups throughout history |  |
| Thecodontosaurus | 1836 | Magnesian Conglomerate (Late Triassic, Rhaetian) | United Kingdom
( England) | Could have been an extremely fast bipedal runner [83] |  |
| Thecospondylus | 1882 | Wealden Group (Early Cretaceous, Valanginian to Hauterivian) | United Kingdom
( England) | Indeterminate within Dinosauria |  |
| Torvosaurus | 1979 | Kimmeridge Clay?, Lourinhã Formation, Ornatenton Formation, Vega Formation? (Middle Jurassic to Late Jurassic, Callovian to Tithonian) | Germany
Portugal
Spain?
United Kingdom
( England?) | The type species was found in the United States. Several species lived around the world, including in Europe, but T. gurneyi is the only non-North American species to receive a formal name. It represents Europe's largest terrestrial predator [84] |  |
| Transylvanosaurus | 2022 | Pui Beds (Late Cretaceous, Maastrichtian) | Romania | Had an unusually wide skull compared to other rhabdodontids |  |
| Trimucrodon | 1973 | Lourinhã Formation (Late Jurassic, Kimmeridgian) | Portugal | Possessed wedge-shaped teeth | |
| Tuebingosaurus | 2022 | Trossingen Formation (Late Triassic, Norian) | Germany | Although originally assigned to Plateosaurus , it contains several features that point to a more derived position |  |
| Turiasaurus | 2006 | Villar del Arzobispo Formation (Late Jurassic, Kimmeridgian to Tithonian) | Spain | Extremely large despite not being a member of Neosauropoda |  |
| Uriash | 2025 | Densuș-Ciula Formation (Late Cretaceous, Maastrichtian) | Romania | One of the largest European sauropods |  |
| Valdosaurus | 1977 | Weald Clay, Wealden Group, Wessex Formation (Early Cretaceous, Berriasian to Barremian) | United Kingdom
( England) | Large and similar to Dryosaurus |  |
| Vallibonavenatrix | 2019 | Arcillas de Morella Formation (Early Cretaceous, Barremian) | Spain | One of the most complete spinosaurids known from Iberia |  |
| Variraptor | 1998 | Argiles et Grès à Reptiles Formation (Late Cretaceous, Campanian to Maastrichtian) | France | May be closely related to Bambiraptor [68] |  |
| Vectaerovenator | 2020 | Lower Greensand Group (Early Cretaceous, Aptian) | United Kingdom
( England) | Only known from four vertebrae but are distinct enough to be classified as their own genus |  |
| Vectidromeus | 2024 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Originally described as a relative of the coeval Hypsilophodon , but a subsequent analysis suggests possible dryosaurid affinities for this genus [85] |  |
| Vectipelta | 2023 | Wessex Formation (Early Cretaceous, Hauterivian to Barremian) | United Kingdom
( England) | May have been more closely related to Asian ankylosaurs than to European ones [86] |  |
| Vectiraptor | 2021 | Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Shares some features with North American dromaeosaurids |  |
| Velocipes | 1932 | Lissauer Breccia (Late Triassic, Norian) |  Poland Poland | Has been considered a dubious, indeterminate vertebrate [87] |  |
| Volgatitan | 2018 | Unnamed formation (Early Cretaceous, Hauterivian) |  Russia Russia | Closely related to South American titanosaurs |  |
| Vouivria | 2017 | Calcaires de Clerval (Late Jurassic, Oxfordian) | France | The oldest known titanosauriform |  |
| Wiehenvenator | 2016 | Ornatenton Formation (Middle Jurassic, Callovian) | Germany | Before its formal description, it had been nicknamed the "Minden Monster" |  |
| Xenoposeidon | 2007 | Wealden Group (Early Cretaceous, Berriasian to Valanginian) | United Kingdom
( England) | Only known from a single, very unique vertebra | 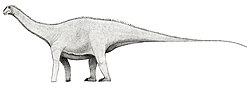 |
| Yaverlandia | 1971 | Vectis Formation?, Wessex Formation (Early Cretaceous, Barremian) | United Kingdom
( England) | Originally misidentified as a pachycephalosaur because of its thick skull roof |  |
| Zalmoxes | 2003 | Densuș-Ciula Formation, Sânpetru Formation, Sebeș Formation (Late Cretaceous, Maastrichtian) | Romania | Has been suggested to be a European ceratopsian [88] |  |
| Zby | 2014 | Lourinhã Formation (Late Jurassic, Kimmeridgian) | Portugal | Originally believed to be a specimen of Turiasaurus |  |
|






















































































































































































































