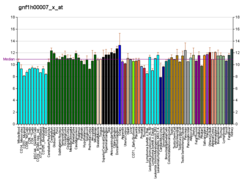
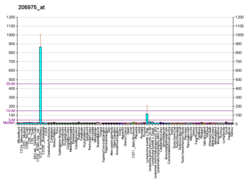
Cytokines are a broad and loose category of small proteins important in cell signaling. Due to their size, cytokines cannot cross the lipid bilayer of cells to enter the cytoplasm and therefore typically exert their functions by interacting with specific cytokine receptors on the target cell surface. Cytokines have been shown to be involved in autocrine, paracrine and endocrine signaling as immunomodulating agents.

Tumor necrosis factor ('TNF), also known as cachexin or cachectin, is a cytokine and a member of the TNF superfamily, which comprises various transmembrane proteins with a homologous TNF domain. Initially referred to as TNFα, this cytokine was the first to be identified as an adipokine, secreted by adipose tissue. TNF signaling occurs through two receptors: TNFR1 and TNFR2. TNFR1 is widely expressed across most cell types and is typically associated with pro-inflammatory and apoptotic signaling. In contrast, TNFR2 is mainly found on endothelial, epithelial, and immune cells, where it mediates anti-inflammatory responses and promotes cell proliferation.

Interleukin 12 (IL-12) is an interleukin that is naturally produced by dendritic cells, macrophages, neutrophils, helper T cells and human B-lymphoblastoid cells (NC-37) in response to antigenic stimulation. IL-12 belongs to the family of interleukin-12. IL-12 family is unique in comprising the only heterodimeric cytokines, which includes IL-12, IL-23, IL-27 and IL-35. Despite sharing many structural features and molecular partners, they mediate surprisingly diverse functional effects.

Transforming growth factor beta (TGF-β) is a multifunctional cytokine belonging to the transforming growth factor superfamily that includes three different mammalian isoforms and many other signaling proteins. TGFB proteins are produced by all white blood cell lineages.

The tumor necrosis factor receptor superfamily (TNFRSF) is a protein superfamily of cytokine receptors characterized by the ability to bind tumor necrosis factors (TNFs) via an extracellular cysteine-rich domain. With the exception of nerve growth factor (NGF), all TNFs are homologous to the archetypal TNF-alpha. In their active form, the majority of TNF receptors form trimeric complexes in the plasma membrane. Accordingly, most TNF receptors contain transmembrane domains (TMDs), although some can be cleaved into soluble forms, and some lack a TMD entirely. In addition, most TNF receptors require specific adaptor protein such as TRADD, TRAF, RIP and FADD for downstream signalling. TNF receptors are primarily involved in apoptosis and inflammation, but they can also take part in other signal transduction pathways, such as proliferation, survival, and differentiation. TNF receptors are expressed in a wide variety of tissues in mammals, especially in leukocytes.
Lymphotoxin is a member of the tumor necrosis factor (TNF) superfamily of cytokines, whose members are responsible for regulating the growth and function of lymphocytes and are expressed by a wide variety of cells in the body.

Tumor necrosis factor receptor superfamily, member 4 (TNFRSF4), also known as CD134 and OX40 receptor, is a member of the TNFR-superfamily of receptors which is not constitutively expressed on resting naïve T cells, unlike CD28. OX40 is a secondary co-stimulatory immune checkpoint molecule, expressed after 24 to 72 hours following activation; its ligand, OX40L, is also not expressed on resting antigen presenting cells, but is following their activation. Expression of OX40 is dependent on full activation of the T cell; without CD28, expression of OX40 is delayed and of fourfold lower levels.

TNF receptor-associated factor 2 is a protein that in humans is encoded by the TRAF2 gene.

Lymphotoxin beta receptor (LTBR), also known as tumor necrosis factor receptor superfamily member 3 (TNFRSF3), is a cell surface receptor for lymphotoxin involved in apoptosis and cytokine release. It is a member of the tumor necrosis factor receptor superfamily.
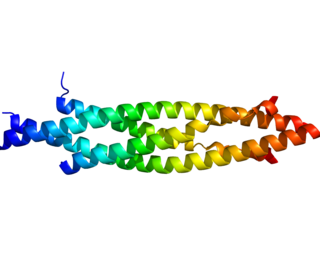
IKK-β also known as inhibitor of nuclear factor kappa-B kinase subunit beta is a protein that in humans is encoded by the IKBKB gene.

TNF receptor-associated factor 1 is a protein that in humans is encoded by the TRAF1 gene.

TNF receptor-associated factor (TRAF3) is a protein that in humans is encoded by the TRAF3 gene.

Lymphotoxin-beta (LT-beta) also known as tumor necrosis factor C (TNF-C) is a protein that in humans is encoded by the LTB gene.
Interferon alpha-1 is a protein that in humans is encoded by the IFNA1 gene.

Mitogen-activated protein kinase kinase kinase 14 (MAP3K14), also known as NF-kappa-B-inducing kinase (NIK), is a MAP kinase kinase kinase enzyme that in humans is encoded by the MAP3K14 gene.

LIGHT, also known as tumor necrosis factor superfamily member 14 (TNFSF14), is a secreted protein of the TNF superfamily. It is recognized by herpesvirus entry mediator (HVEM), as well as decoy receptor 3.

Tumor necrosis factor receptor superfamily member 18 (TNFRSF18), also known as glucocorticoid-induced TNFR-related protein (GITR) or CD357. GITR is encoded and tnfrsf18 gene at chromosome 4 in mice. GITR is type I transmembrane protein and is described in 4 different isoforms. GITR human orthologue, also called activation-inducible TNFR family receptor (AITR), is encoded by the TNFRSF18 gene at chromosome 1.
The following outline is provided as an overview of and topical guide to immunology:
Innate lymphoid cells (ILCs) are the most recently discovered family of innate immune cells, derived from common lymphoid progenitors (CLPs). In response to pathogenic tissue damage, ILCs contribute to immunity via the secretion of signalling molecules, and the regulation of both innate and adaptive immune cells. ILCs are primarily tissue resident cells, found in both lymphoid, and non- lymphoid tissues, and rarely in the blood. They are particularly abundant at mucosal surfaces, playing a key role in mucosal immunity and homeostasis. Characteristics allowing their differentiation from other immune cells include the regular lymphoid morphology, absence of rearranged antigen receptors found on T cells and B cells, and phenotypic markers usually present on myeloid or dendritic cells.
Immunology is the study of the immune system during health and disease. Below is a list of immunology-related articles.