Free-running sleep is a sleep pattern that is not adjusted (entrained) to the 24-hour cycle in nature nor to any artificial cycle.

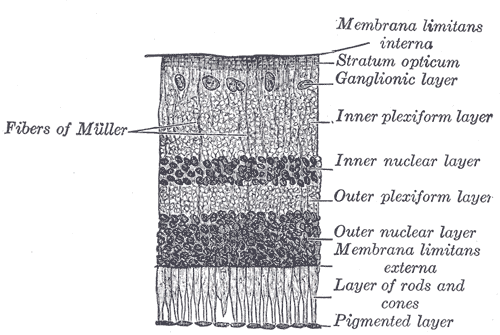
The retina is the innermost, light-sensitive layer of tissue of the eye of most vertebrates and some molluscs. The optics of the eye create a focused two-dimensional image of the visual world on the retina, which translates that image into electrical neural impulses to the brain to create visual perception. The retina serves a function analogous to that of the film or image sensor in a camera.

The visual system comprises the sensory organ and parts of the central nervous system which gives organisms the sense of sight as well as enabling the formation of several non-image photo response functions. It detects and interprets information from the optical spectrum perceptible to that species to "build a representation" of the surrounding environment. The visual system carries out a number of complex tasks, including the reception of light and the formation of monocular neural representations, colour vision, the neural mechanisms underlying stereopsis and assessment of distances to and between objects, the identification of particular object of interest, motion perception, the analysis and integration of visual information, pattern recognition, accurate motor coordination under visual guidance, and more. The neuropsychological side of visual information processing is known as visual perception, an abnormality of which is called visual impairment, and a complete absence of which is called blindness. Non-image forming visual functions, independent of visual perception, include the pupillary light reflex (PLR) and circadian photoentrainment.

A photoreceptor cell is a specialized type of neuroepithelial cell found in the retina that is capable of visual phototransduction. The great biological importance of photoreceptors is that they convert light into signals that can stimulate biological processes. To be more specific, photoreceptor proteins in the cell absorb photons, triggering a change in the cell's membrane potential.

Rod cells are photoreceptor cells in the retina of the eye that can function in lower light better than the other type of visual photoreceptor, cone cells. Rods are usually found concentrated at the outer edges of the retina and are used in peripheral vision. On average, there are approximately 92 million rod cells in the human retina. Rod cells are more sensitive than cone cells and are almost entirely responsible for night vision. However, rods have little role in color vision, which is the main reason why colors are much less apparent in dim light.

A retinal ganglion cell (RGC) is a type of neuron located near the inner surface of the retina of the eye. It receives visual information from photoreceptors via two intermediate neuron types: bipolar cells and retina amacrine cells. Retina amacrine cells, particularly narrow field cells, are important for creating functional subunits within the ganglion cell layer and making it so that ganglion cells can observe a small dot moving a small distance. Retinal ganglion cells collectively transmit image-forming and non-image forming visual information from the retina in the form of action potential to several regions in the thalamus, hypothalamus, and mesencephalon, or midbrain.
Miosis, or myosis, is excessive constriction of the pupil. The term is from Ancient Greek μύειν mūein, "to close the eyes". The opposite condition, mydriasis, is the dilation of the pupil. Anisocoria is the condition of one pupil being more dilated than the other.

Melanopsin is a type of photopigment belonging to a larger family of light-sensitive retinal proteins called opsins and encoded by the gene Opn4. In the mammalian retina, there are two additional categories of opsins, both involved in the formation of visual images: rhodopsin and photopsin in the rod and cone photoreceptor cells, respectively.

Opsins are a group of proteins, made light-sensitive, via the chromophore retinal found in photoreceptor cells of the retina. Five classical groups of opsins are involved in vision, mediating the conversion of a photon of light into an electrochemical signal, the first step in the visual transduction cascade. Another opsin found in the mammalian retina, melanopsin, is involved in circadian rhythms and pupillary reflex but not in vision.

Giant retinal ganglion cells are photosensitive ganglion cells with large dendritic trees discovered in the human and macaque retina by Dacey et al. (2005).
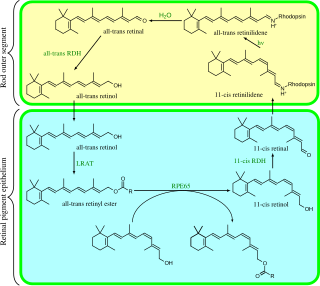
Visual phototransduction is the sensory transduction of the visual system. It is a process by which light is converted into electrical signals in the rod cells, cone cells and photosensitive ganglion cells of the retina of the eye. This cycle was elucidated by George Wald (1906–1997) for which he received the Nobel Prize in 1967. It is so called "Wald's Visual Cycle" after him.
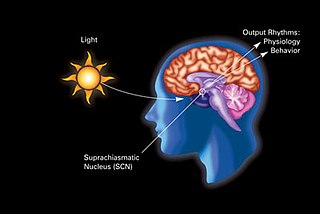
The retinohypothalamic tract (RHT) is a photic neural input pathway involved in the circadian rhythms of mammals. The origin of the retinohypothalamic tract is the intrinsically photosensitive retinal ganglion cells (ipRGC), which contain the photopigment melanopsin. The axons of the ipRGCs belonging to the retinohypothalamic tract project directly, monosynaptically, to the suprachiasmatic nuclei (SCN) via the optic nerve and the optic chiasm. The suprachiasmatic nuclei receive and interpret information on environmental light, dark and day length, important in the entrainment of the "body clock". They can coordinate peripheral "clocks" and direct the pineal gland to secrete the hormone melatonin.
Ignacio Provencio is an American neuroscientist and the discoverer of melanopsin, a photopigment found in specialized photosensitive ganglion cells of the mammalian retina. Provencio served as the program committee chair of the Society for Research on Biological Rhythms from 2008 to 2010.
Photoreceptor proteins are light-sensitive proteins involved in the sensing and response to light in a variety of organisms. Some examples are rhodopsin in the photoreceptor cells of the vertebrate retina, phytochrome in plants, and bacteriorhodopsin and bacteriophytochromes in some bacteria. They mediate light responses as varied as visual perception, phototropism and phototaxis, as well as responses to light-dark cycles such as circadian rhythm and other photoperiodisms including control of flowering times in plants and mating seasons in animals.
Light effects on circadian rhythm are the effects that light has on circadian rhythm.
Mammals normally have a pair of eyes. Although mammalian vision is not so excellent as bird vision, it is at least dichromatic for most of mammalian species, with certain families possessing a trichromatic color perception.

Russell Grant Foster, CBE, FRS FMedSci is a British professor of circadian neuroscience, the Director of the Nuffield Laboratory of Ophthalmology and the Head of the Sleep and Circadian Neuroscience Institute (SCNi). He is also a Nicholas Kurti Senior Fellow at the Brasenose College at the University of Oxford. Foster and his group are credited with key contributions to the discovery of the non-rod, non-cone, photosensitive retinal ganglion cells (pRGCs) in the mammalian retina which provide input to the circadian rhythm system. He has written and co-authored over a hundred scientific publications.

A midget cell is one type of retinal ganglion cell (RGC). Midget cells originate in the ganglion cell layer of the retina, and project to the parvocellular layers of the lateral geniculate nucleus (LGN). The axons of midget cells travel through the optic nerve and optic tract, ultimately synapsing with parvocellular cells in the LGN. These cells are known as midget retinal ganglion cells due to the small sizes of their dendritic trees and cell bodies. About 80% of RGCs are midget cells. They receive inputs from relatively few rods and cones. In many cases, they are connected to midget bipolar cells, which are linked to one cone each.

King-Wai Yau is a Chinese-born American neuroscientist and Professor of Neuroscience at Johns Hopkins University School of Medicine in Baltimore, Maryland.
Samer Hattar is a chronobiologist and a leader in the field of non-image forming photoreception. He is currently the Chief of the Section on Light and Circadian Rhythms at the National Institute of Mental Health, part of the National Institutes of Health. He was previously an associate professor in the Department of Neuroscience and the Department of Biology at Johns Hopkins University in Baltimore, MD. He is best known for his investigation into the role of melanopsin and intrinsically photosensitive retinal ganglion cells (ipRGC) in the entrainment of circadian rhythms.