
Titanosaurs were a diverse group of sauropod dinosaurs, including genera from all seven continents. The titanosaurs were the last surviving group of long-necked sauropods, with taxa still thriving at the time of the extinction event at the end of the Cretaceous. This group includes some of the largest land animals known to have ever existed, such as Patagotitan, estimated at 37 m (121 ft) long with a weight of 69 tonnes, and the comparably-sized Argentinosaurus and Puertasaurus from the same region.

Bonitasaura is a genus of titanosaurian dinosaur hailing from uppermost layers of the Late Cretaceous (Santonian) Bajo de la Carpa Formation, Neuquén Group of the eastern Neuquén Basin, located in Río Negro Province, Northwestern Patagonia, Argentina. The remains, consisting of a partial sub-adult skeleton jumbled in a small area of fluvial sandstone, including a lower jaw with teeth, a partial vertebrae series, and limb bones, were described by Sebastian Apesteguía in 2004.

Saichania is a genus of herbivorous ankylosaurid dinosaur from the Late Cretaceous period of Mongolia and China.

Ampelosaurus is a titanosaurian sauropod dinosaur from the Late Cretaceous Period of what is now France. Its type species is A. atacis, named by Le Loeuff in 1995. Its remains were found in a level dating from 71.5 million years ago representing the early Maastrichtian.

Andesaurus is a genus of basal titanosaurian sauropod dinosaur which existed during the middle of the Cretaceous Period in South America. Like most sauropods, belonging to one of the largest animals ever to walk the Earth, it would have had a small head on the end of a long neck and an equally long tail.

Epachthosaurus was a genus of titanosaurian sauropod dinosaur from the Late Cretaceous. It was a basal lithostrotian titanosaur. Its fossils have been found in Central and Northern Patagonia in South America.

Jiangjunosaurus is a genus of herbivorous stegosaurian dinosaur from the Oxfordian-age Shishugou Formation of the Junggar Basin, Xinjiang, China.

Miragaia is a long-necked stegosaurid dinosaur. Its fossils have been found in Upper Jurassic rocks in Portugal and possibly also Wyoming, United States. Miragaia has the longest neck known for any stegosaurian, which included at least seventeen vertebrae. Some researchers consider this taxon to be a junior synonym of Dacentrurus.

Jesairosaurus is an extinct genus of early archosauromorph reptile known from the Illizi Province of Algeria. It is known from a single species, Jesairosaurus lehmani. Although a potential relative of the long-necked tanystropheids, this lightly-built reptile could instead be characterized by its relatively short neck as well as various skull features.
This Glossary explains technical terms commonly employed in the description of dinosaur body fossils. Besides dinosaur-specific terms, it covers terms with wider usage, when these are of central importance in the study of dinosaurs or when their discussion in the context of dinosaurs is beneficial. The glossary does not cover ichnological and bone histological terms, nor does it cover measurements.

Yongjinglong is an extinct genus of titanosauriform sauropod dinosaur known from the Early Cretaceous of Lanzhou-Minhe Basin of Gansu Province, China. It contains a single species, Yongjinglong datangi.
Patagotitan is a genus of titanosaurian sauropod dinosaur from the Cerro Barcino Formation in Chubut Province, Patagonia, Argentina. The genus contains a single species known from at least six young adult individuals, Patagotitan mayorum, which was first announced in 2014 and then named in 2017 by José Carballido and colleagues. Preliminary studies and press releases suggested that Patagotitan was the largest known titanosaur and land animal overall, with an estimated length of 37 m (121 ft) and an estimated weight of 69 tonnes. Later research revised the length estimate down to 31 m (102 ft) and weight estimates down to approximately 50–57 tonnes, suggesting that Patagotitan was of a similar size to, if not smaller than, its closest relatives Argentinosaurus and Puertasaurus. Still, Patagotitan is one of the most-known titanosaurs, and so its interrelationships with other titanosaurs have been relatively consistent in phylogenetic analyses. This led to its use in a re-definition of the group Colossosauria by Carballido and colleagues in 2022.

Dreadnoughtus is a genus of titanosaurian sauropod dinosaur containing a single species, Dreadnoughtus schrani. D. schrani is known from two partial skeletons discovered in Upper Cretaceous rocks of the Cerro Fortaleza Formation in Santa Cruz Province, Argentina. It is one of the largest terrestrial vertebrates known, with the immature type specimen measuring 26 metres (85 ft) in total body length and weighing 48–49 metric tons. D. schrani is known from more complete skeletons than any other gigantic titanosaurian.
Lohuecotitan is an extinct genus of titanosaurian sauropod dinosaur which lived during the Late Cretaceous in Spain. The only species known in the genus is Lohuecotitan pandafilandi, described and named in 2016.
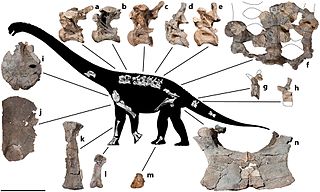
Savannasaurus is a genus of titanosaurian sauropod dinosaur from the Late Cretaceous Winton Formation of Queensland, Australia. It contains one species, Savannasaurus elliottorum, named in 2016 by Stephen Poropat and colleagues. The holotype and only known specimen, originally nicknamed "Wade", is the most complete specimen of an Australian sauropod, and is held at the Australian Age of Dinosaurs museum. Dinosaurs known from contemporary rocks include its close relative Diamantinasaurus and the theropod Australovenator; associated teeth suggest that Australovenator may have fed on the holotype specimen.
Magnamanus is an extinct genus of herbivorous iguanodontian dinosaur that lived during the Early Cretaceous in what is now Spain in the Golmayo Formation. It contains a single species, Magnamanus soriaensis.

Mansourasaurus is a genus of herbivorous lithostrotian sauropod dinosaur from the Quseir Formation of Egypt. The type and only species is Mansourasaurus shahinae.

Mnyamawamtuka is a genus of lithostrotian titanosaur sauropod dinosaur from the Cretaceous Galula Formation in Tanzania. The type and only species is M. moyowamkia.
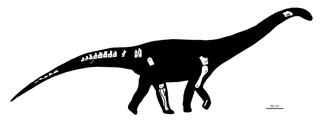
Nullotitan is a genus of lithostrotian titanosaur from the Chorrillo Formation from Santa Cruz Province in Argentina. The type and only species is Nullotitan glaciaris. It was a contemporary of the ornithopod Isasicursor which was described in the same paper.

Stegouros is a genus of ankylosaurian dinosaur from the Late Cretaceous Dorotea Formation of southern Chile. The genus contains a single species, Stegouros elengassen, known from a semi-articulated, near-complete skeleton.
















