Related Research Articles

Ecology is the natural science of the relationships among living organisms, including humans, and their physical environment. Ecology considers organisms at the individual, population, community, ecosystem, and biosphere levels. Ecology overlaps with the closely related sciences of biogeography, evolutionary biology, genetics, ethology, and natural history.

Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.

In ecology, a niche is the match of a species to a specific environmental condition. It describes how an organism or population responds to the distribution of resources and competitors and how it in turn alters those same factors. "The type and number of variables comprising the dimensions of an environmental niche vary from one species to another [and] the relative importance of particular environmental variables for a species may vary according to the geographic and biotic contexts".

Biological dispersal refers to both the movement of individuals from their birth site to their breeding site and the movement from one breeding site to another . Dispersal is also used to describe the movement of propagules such as seeds and spores. Technically, dispersal is defined as any movement that has the potential to lead to gene flow. The act of dispersal involves three phases: departure, transfer, and settlement. There are different fitness costs and benefits associated with each of these phases. Through simply moving from one habitat patch to another, the dispersal of an individual has consequences not only for individual fitness, but also for population dynamics, population genetics, and species distribution. Understanding dispersal and the consequences, both for evolutionary strategies at a species level and for processes at an ecosystem level, requires understanding on the type of dispersal, the dispersal range of a given species, and the dispersal mechanisms involved. Biological dispersal can be correlated to population density. The range of variations of a species' location determines the expansion range.

Population ecology is a sub-field of ecology that deals with the dynamics of species populations and how these populations interact with the environment, such as birth and death rates, and by immigration and emigration.

Habitat fragmentation describes the emergence of discontinuities (fragmentation) in an organism's preferred environment (habitat), causing population fragmentation and ecosystem decay. Causes of habitat fragmentation include geological processes that slowly alter the layout of the physical environment, and human activity such as land conversion, which can alter the environment much faster and causes the extinction of many species. More specifically, habitat fragmentation is a process by which large and contiguous habitats get divided into smaller, isolated patches of habitats.

A metapopulation consists of a group of spatially separated populations of the same species which interact at some level. The term metapopulation was coined by Richard Levins in 1969 to describe a model of population dynamics of insect pests in agricultural fields, but the idea has been most broadly applied to species in naturally or artificially fragmented habitats. In Levins' own words, it consists of "a population of populations".
Evolutionary ecology lies at the intersection of ecology and evolutionary biology. It approaches the study of ecology in a way that explicitly considers the evolutionary histories of species and the interactions between them. Conversely, it can be seen as an approach to the study of evolution that incorporates an understanding of the interactions between the species under consideration. The main subfields of evolutionary ecology are life history evolution, sociobiology, the evolution of interspecific interactions and the evolution of biodiversity and of ecological communities.

Molecular ecology is a subdiscipline of ecology that is concerned with applying molecular genetic techniques to ecological questions. It is virtually synonymous with the field of "Ecological Genetics" as pioneered by Theodosius Dobzhansky, E. B. Ford, Godfrey M. Hewitt, and others. Molecular ecology is related to the fields of population genetics and conservation genetics.
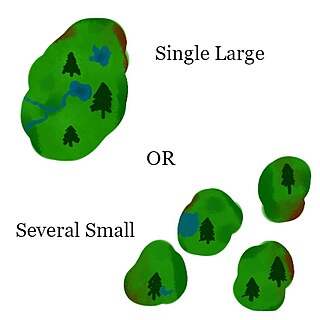
The SLOSS debate was a debate in ecology and conservation biology during the 1970's and 1980's as to whether a single large or several small (SLOSS) reserves were a superior means of conserving biodiversity in a fragmented habitat. Since its inception, multiple alternate theories have been proposed. There have been applications of the concept outside of the original context of habitat conservation.
In landscape ecology, landscape connectivity is, broadly, "the degree to which the landscape facilitates or impedes movement among resource patches". Alternatively, connectivity may be a continuous property of the landscape and independent of patches and paths. Connectivity includes both structural connectivity and functional connectivity. Functional connectivity includes actual connectivity and potential connectivity in which movement paths are estimated using the life-history data.

Interspecific competition, in ecology, is a form of competition in which individuals of different species compete for the same resources in an ecosystem. This can be contrasted with mutualism, a type of symbiosis. Competition between members of the same species is called intraspecific competition.

In ecology, a community is a group or association of populations of two or more different species occupying the same geographical area at the same time, also known as a biocoenosis, biotic community, biological community, ecological community, or life assemblage. The term community has a variety of uses. In its simplest form it refers to groups of organisms in a specific place or time, for example, "the fish community of Lake Ontario before industrialization".

Cross-boundary subsidies are caused by organisms or materials that cross or traverse habitat patch boundaries, subsidizing the resident populations. The transferred organisms and materials may provide additional predators, prey, or nutrients to resident species, which can affect community and food web structure. Cross-boundary subsidies of materials and organisms occur in landscapes composed of different habitat patch types, and so depend on characteristics of those patches and on the boundaries in between them. Human alteration of the landscape, primarily through fragmentation, has the potential to alter important cross-boundary subsidies to increasingly isolated habitat patches. Understanding how processes that occur outside of habitat patches can affect populations within them may be important to habitat management.
Extinction threshold is a term used in conservation biology to explain the point at which a species, population or metapopulation, experiences an abrupt change in density or number because of an important parameter, such as habitat loss. It is at this critical value below which a species, population, or metapopulation, will go extinct, though this may take a long time for species just below the critical value, a phenomenon known as extinction debt.
In ecology, the occupancy–abundance (O–A) relationship is the relationship between the abundance of species and the size of their ranges within a region. This relationship is perhaps one of the most well-documented relationships in macroecology, and applies both intra- and interspecifically. In most cases, the O–A relationship is a positive relationship. Although an O–A relationship would be expected, given that a species colonizing a region must pass through the origin and could reach some theoretical maximum abundance and distribution, the relationship described here is somewhat more substantial, in that observed changes in range are associated with greater-than-proportional changes in abundance. Although this relationship appears to be pervasive, and has important implications for the conservation of endangered species, the mechanism(s) underlying it remain poorly understood.
Ecological traps are scenarios in which rapid environmental change leads organisms to prefer to settle in poor-quality habitats. The concept stems from the idea that organisms that are actively selecting habitat must rely on environmental cues to help them identify high-quality habitat. If either the habitat quality or the cue changes so that one does not reliably indicate the other, organisms may be lured into poor-quality habitat.
An ecological metacommunity is a set of interacting communities which are linked by the dispersal of multiple, potentially interacting species. The term is derived from the field of community ecology, which is primarily concerned with patterns of species distribution, abundance and interactions. Metacommunity ecology combines the importance of local factors and regional factors to explain patterns of species distributions that happen in different spatial scales.
In macroecology and community ecology, an occupancy frequency distribution (OFD) is the distribution of the numbers of species occupying different numbers of areas. It was first reported in 1918 by the Danish botanist Christen C. Raunkiær in his study on plant communities. The OFD is also known as the species-range size distribution in literature.
The rescue effect is a phenomenon which was first described by Brown and Kodric-Brown, and is commonly used in metapopulation dynamics and many other disciplines in ecology. This populational process explains how the migration of individuals can increase the persistence of small isolated populations by helping to stabilize a metapopulation, thus reducing the chances of extinction. In other words, immigration can lead to the recolonization of previously extinct patches, promoting the long-term persistence of the network of populations.
References
- ↑ Holt RD (October 1985). "Population dynamics in two-patch environments: some anomalous consequences of an optimal habitat distribution". Theoretical Population Biology. 28 (2): 181–208. doi:10.1016/0040-5809(85)90027-9.
- 1 2 3 4 Pulliam HR (November 1988). "Sources, sinks, and population regulation". The American Naturalist. 132 (5): 652–61. doi:10.1086/284880. S2CID 84423952.
- 1 2 Watkinson AR, Sutherland WJ (January 1995). "Sources, sinks and pseudo-sinks". Journal of Animal Ecology. 1 (1): 126–30. Bibcode:1995JAnEc..64..126W. doi:10.2307/5833. JSTOR 5833.
- 1 2 Thomas CD, Singer MC, Boughton DA (December 1996). "Catastrophic extinction of population sources in a butterfly metapopulation". The American Naturalist. 148 (6): 957–75. doi:10.1086/285966. S2CID 85253063.
- 1 2 Dias PC (August 1996). "Sources and sinks in population biology". Trends in Ecology & Evolution. 11 (8): 326–30. doi:10.1016/0169-5347(96)10037-9. PMID 21237863.
- ↑ Johnson DM (July 2004). "Source–sink dynamics in a temporally heterogeneous environment". Ecology. 85 (7): 2037–45. Bibcode:2004Ecol...85.2037J. doi:10.1890/03-0508.
- ↑ Boughton DA (December 1999). "Empirical evidence for complex source–sink dynamics with alternative states in a butterfly metapopulation". Ecology. 80 (8): 2727–39. doi:10.1890/0012-9658(1999)080[2727:EEFCSS]2.0.CO;2.
- 1 2 Tittler R, Fahrig L, Villard MA (December 2006). "Evidence of large-scale source-sink dynamics and long-distance dispersal among Wood Thrush populations". Ecology. 87 (12): 3029–36. doi:10.1890/0012-9658(2006)87[3029:eolsda]2.0.co;2. PMID 17249228.
- 1 2 3 Runge JP, Runge MC, Nichols JD (June 2006). "The role of local populations within a landscape context: defining and classifying sources and sinks". The American Naturalist. 167 (6): 925–38. doi:10.1086/503531. PMID 16615034. S2CID 8952958.
- ↑ Caswell H (2018). Matrix population models : construction, analysis, and interpretation (2nd ed.). Sunderland, Mass.: Sinauer Associates. ISBN 978-0-87893-096-8.
- ↑ Williams BK, Nichols JD, Conroy MJ (2002). Analysis and management of animal populations : modeling, estimation, and decision making. San Diego: Academic Press. ISBN 978-0-08-057472-1.
- ↑ Keddy PA (January 1982). "Population ecology on an environmental gradient: Cakile edentula on a sand dune". Oecologia. 52 (3): 348–355. Bibcode:1982Oecol..52..348K. doi:10.1007/BF00367958. PMID 28310394. S2CID 7778352.
- ↑ Foissner W (1987). "Soil protozoa: fundamental problems, ecological significance, adaptations in ciliates and testaceans, bioindicators, and guide to the literature". Progress in Protistology. 2: 69–212.
- ↑ Fernández LD (February 2015). "Source–sink dynamics shapes the spatial distribution of soil protists in an arid shrubland of northern Chile". Journal of Arid Environments. 113: 121–5. Bibcode:2015JArEn.113..121F. doi:10.1016/j.jaridenv.2014.10.007.
- 1 2 3 Diffendorfer JE (April 1998). "Testing models of source-sink dynamics and balanced dispersal". Oikos. 81 (3): 417–33. Bibcode:1998Oikos..81..417D. doi:10.2307/3546763. JSTOR 3546763.
- ↑ Fretwell SD, Lucas Jr HL (March 1969). "On territorial behavior and other factors influencing habitat distribution in birds". Acta Biotheoretica. 19 (1): 45–52. doi:10.1007/BF01601955. S2CID 89682949.
- ↑ Pulliam HR, Danielson BJ (June 1991). "Sources, sinks, and habitat selection: a landscape perspective on population dynamics". The American Naturalist. 137: S50-66. doi:10.1086/285139. S2CID 85125604.
- ↑ Grinnell J (October 1917). "The niche-relationships of the California Thrasher". The Auk. 34 (4): 427–33. doi:10.2307/4072271. JSTOR 4072271.
- ↑ Hutchinson GE (1957). Concluding remarks. Cold Spring Harbor Symposium Quantitative Biology. Vol. 22. pp. 415–427.
- ↑ Manlik O, Chabanne D, Daniel C, Bejder L, Allen SJ, Sherwin WB (2018). "Demography and genetics suggest reversal of dolphin source-sink dynamics, with implications for conservation". Marine Mammal Science. 35 (3): 732–759. doi:10.1111/mms.12555. S2CID 92108810.
- ↑ Howe RW, Davis GJ, Mosca V (January 1991). "The demographic significance of 'sink'populations". Biological Conservation. 57 (3): 239–55. Bibcode:1991BCons..57..239H. doi:10.1016/0006-3207(91)90071-G.
- ↑ Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, et al. (July 2004). "The metacommunity concept: a framework for multi-scale community ecology". Ecology Letters. 7 (7): 601–13. Bibcode:2004EcolL...7..601L. doi: 10.1111/j.1461-0248.2004.00608.x .
- ↑ Robertson BA, Hutto RL (May 2006). "A framework for understanding ecological traps and an evaluation of existing evidence". Ecology. 87 (5): 1075–85. doi:10.1890/0012-9658(2006)87[1075:AFFUET]2.0.CO;2. PMID 16761584. S2CID 266029513.
- ↑ Frouz J, Kindlmann P (2001). "The role of sink to source re-colonisation in the population dynamics of insects living in unstable habitats: an example of terrestrial chironomids". Oikos. 93 (1): 50–58. Bibcode:2001Oikos..93...50F. doi:10.1034/j.1600-0706.2001.930105.x. ISSN 1600-0706.
- ↑ Frouz J, Kindlmann P (2015-06-05). "Source-Sink Colonization as a Possible Strategy of Insects Living in Temporary Habitats". PLOS ONE. 10 (6): e0127743. Bibcode:2015PLoSO..1027743F. doi: 10.1371/journal.pone.0127743 . PMC 4457784 . PMID 26047010.
- ↑ Lamb CT, Mowat G, McLellan BN, Nielsen SE, Boutin S (January 2017). "Forbidden fruit: human settlement and abundant fruit create an ecological trap for an apex omnivore". The Journal of Animal Ecology. 86 (1): 55–65. Bibcode:2017JAnEc..86...55L. doi: 10.1111/1365-2656.12589 . PMID 27677529.
- 1 2 3 Roberts CM. "Sources, sinks, and the design of marine reserve networks". Fisheries. 23: 16–19.
- ↑ Lee DE, Bolger DT (2017-05-05). "Movements and source–sink dynamics of a Masai giraffe metapopulation". Population Ecology. 59 (2): 157–168. Bibcode:2017PopEc..59..157L. doi:10.1007/s10144-017-0580-7. ISSN 1438-3896. S2CID 24276021.