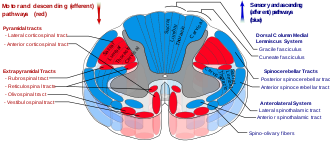
Tracts
The main spinocerebellar tracts are the dorsal and the ventral spinocerebellar tracts. [1]
Ventral spinocerebellar tract
The ventral spinocerebellar tract (or anterior spinocerebellar tract) conveys proprioceptive information from the body to the cerebellum. Historically, it has also been known as Gowers' column (or fasciculus or tract), after Sir William Richard Gowers.
It is part of the somatosensory system and runs in parallel with the dorsal spinocerebellar tract. Both these tracts involve two neurons. The ventral spinocerebellar tract will cross to the opposite side of the body first in the spinal cord as part of the anterior white commissure and then cross again to end in the cerebellum (referred to as a "double cross"), as compared to the dorsal spinocerebellar tract, which does not decussate, or cross sides, at all through its path.
The ventral tract (under L2/L3) gets its proprioceptive/fine touch/vibration information from a first order neuron, with its cell body in a dorsal ganglion. The axon runs via the fila radicularia to the dorsal horn of the grey matter. There it makes a synapse with the dendrites of two neurons: they send their axons bilaterally to the ventral border of the lateral funiculi. The fibers of the ventral spinocerebellar tract then enters the cerebellum via the superior cerebellar peduncle. This is in contrast with the dorsal spinocerebellar tract (C8 - L2/L3), which only has 1 unilateral axon that has its cell body in Clarke's column (only at the level of C8 - L2/L3).
Originates from ventral horn at lumbosacral spinal levels. Axons first cross midline in the spinal cord and run in the ventral border of the lateral funiculi. These axons ascend to the pons where they join the superior cerebellar peduncle to enter the cerebellum. Once in the deep white matter of the cerebellum, the axons recross the midline, give off collaterals to the globose and emboliform nuclei, and terminate in the cortex of the anterior lobe and vermis of the posterior lobe.
Comparison with dorsal spinocerebellar tract
When the dorsal roots are cut in a cat performing a step cycle, peripheral excitation is lost, and the dorsal spinocerebellar tract has no activity; the ventral spinocerebellar tract continues to show activity. This suggests that the dorsal spinocerebellar tract carries sensory information to the spinocerebellum through the inferior cerebellar peduncle during movement (since the inferior peduncle is known to contain fibres from the dorsal tract), and that the ventral spinocerebellar tract carries internally generated motor information about the movement through the superior cerebellar peduncle. [4]
Posterior external arcuate fibers
The posterior external arcuate fibers (dorsal external arcuate fibers or cuneocerebellar tract) [5] take origin in the accessory cuneate nucleus, and pass to the inferior cerebellar peduncle of the same side. The term "cuneocerebellar tract" is also used to describe exteroceptive and proprioceptive components that take origin in the gracile and cuneate nuclei; they pass to the inferior cerebellar peduncle of the same side. [6]
The posterior external arcuate fibers carry proprioceptive information from the upper limbs and neck. It is an analogue to the dorsal spinocerebellar tract for the upper limbs. [7] In this context, the "cuneo-" derives from the accessory cuneate nucleus, not the cuneate nucleus. (The two nuclei are related in space, but not in function.)
It is uncertain whether fibers are continued directly from the gracile and cuneate fasciculi into the inferior peduncle.
Rostral spinocerebellar tract
The rostral spinocerebellar tract is a tract which transmits information from the golgi tendon organs of the cranial half of the body to the cerebellum. [8] It terminates bilaterally in the anterior lobe of the cerebellum (lower cerebellar peduncle) after travelling ipsilaterally from its origin in the cervical portion of the spinal cord. [9] [10] It reaches the cerebellum partly through the brachium conjunctivum (superior cerebellar peduncle) and partly through the restiform body (inferior cerebellar peduncle). [10]
Pathway for dorsal and spinocuneocerebellar tracts
The sensory neurons synapse in the posterior thoracic nucleus also known as Clarke's nucleus.
This is a column of relay neuron cell bodies within the medial gray matter within the spinal cord in layer VII (just beneath the dorsal horn), specifically between T1-L3. These neurons then send axons up the spinal cord, and project ipsilaterally to medial zones of the cerebellum through the inferior cerebellar peduncle.
Below L3, relevant neurons pass into the gracile fasciculus (usually associated with the dorsal column–medial lemniscus pathway) until L3 where they synapse with the posterior thoracic nucleus (leading to considerable caudal enlargement).
The neurons in the accessory cuneate nucleus have axons leading to the ipsilateral cerebellum via the inferior cerebellar peduncle.
This page is based on this
Wikipedia article Text is available under the
CC BY-SA 4.0 license; additional terms may apply.
Images, videos and audio are available under their respective licenses.