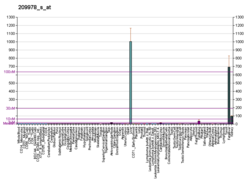
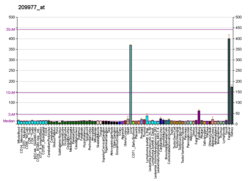
1b2i: KRINGLE 2 DOMAIN OF HUMAN PLASMINOGEN: NMR SOLUTION STRUCTURE OF TRANS-4-AMINOMETHYLCYCLOHEXANE-1-CARBOXYLIC ACID (AMCHA) COMPLEX
1bml: COMPLEX OF THE CATALYTIC DOMAIN OF HUMAN PLASMIN AND STREPTOKINASE
1bui: STRUCTURE OF THE TERNARY MICROPLASMIN-STAPHYLOKINASE-MICROPLASMIN COMPLEX: A PROTEINASE-COFACTOR-SUBSTRATE COMPLEX IN ACTION.
1cea: THE STRUCTURE OF THE NON-COVALENT COMPLEX OF RECOMBINANT KRINGLE 1 DOMAIN OF HUMAN PLASMINOGEN WITH EACA (EPSILON-AMINOCAPROIC ACID)
1ceb: THE STRUCTURE OF THE NON-COVALENT COMPLEX OF RECOMBINANT KRINGLE 1 DOMAIN OF HUMAN PLASMINOGEN WITH AMCHA (TRANS-4-AMINOMETHYLCYCLOHEXANE-1-CARBOXYLIC ACID)
1ddj: CRYSTAL STRUCTURE OF HUMAN PLASMINOGEN CATALYTIC DOMAIN
1hpj: SOLUTION NMR STRUCTURE OF THE HUMAN PLASMINOGEN KRINGLE 1 DOMAIN COMPLEXED WITH 6-AMINOHEXANOIC ACID AT PH 5.3, 310K, DERIVED FROM RANDOMLY GENERATED STRUCTURES USING SIMULATED ANNEALING, 12 STRUCTURES
1hpk: SOLUTION NMR STRUCTURE OF THE HUMAN PLASMINOGEN KRINGLE 1 DOMAIN COMPLEXED WITH 6-AMINOHEXANOIC ACID AT PH 5.3, 310K, DERIVED FROM RANDOMLY GENERATED STRUCTURES USING SIMULATED ANNEALING, MINIMIZED AVERAGE STRUCTURE
1i5k: STRUCTURE AND BINDING DETERMINANTS OF THE RECOMBINANT KRINGLE-2 DOMAIN OF HUMAN PLASMINOGEN TO AN INTERNAL PEPTIDE FROM A GROUP A STREPTOCOCCAL SURFACE PROTEIN
1ki0: The X-ray Structure of Human Angiostatin
1krn: STRUCTURE OF KRINGLE 4 AT 4C TEMPERATURE AND 1.67 ANGSTROMS RESOLUTION
1l4d: CRYSTAL STRUCTURE OF MICROPLASMINOGEN-STREPTOKINASE ALPHA DOMAIN COMPLEX
1l4z: X-RAY CRYSTAL STRUCTURE OF THE COMPLEX OF MICROPLASMINOGEN WITH ALPHA DOMAIN OF STREPTOKINASE IN THE PRESENCE CADMIUM IONS
1pk4: CRYSTAL AND MOLECULAR STRUCTURE OF HUMAN PLASMINOGEN KRINGLE 4 REFINED AT 1.9-ANGSTROMS RESOLUTION
1pkr: THE STRUCTURE OF RECOMBINANT PLASMINOGEN KRINGLE 1 AND THE FIBRIN BINDING SITE
1pmk: KRINGLE-KRINGLE INTERACTIONS IN MULTIMER KRINGLE STRUCTURES
1qrz: CATALYTIC DOMAIN OF PLASMINOGEN
1rjx: Human PLASMINOGEN CATALYTIC DOMAIN, K698M MUTANT
2doh: The X-ray crystallographic structure of the angiogenesis inhibitor, angiostatin, bound a to a peptide from the group A streptococcal surface protein PAM
2doi: The X-ray crystallographic structure of the angiogenesis inhibitor, angiostatin, bound to a peptide from the group A streptococcus protein PAM
2pk4: THE REFINED STRUCTURE OF THE EPSILON-AMINOCAPROIC ACID COMPLEX OF HUMAN PLASMINOGEN KRINGLE
5hpg: STRUCTURE AND LIGAND DETERMINANTS OF THE RECOMBINANT KRINGLE 5 DOMAIN OF HUMAN PLASMINOGEN