Enzyme catalysis
Enzymes have been postulated to use quantum tunneling to transfer electrons in electron transport chains. [13] [14] [15] It is possible that protein quaternary architectures may have adapted to enable sustained quantum entanglement and coherence, which are two of the limiting factors for quantum tunneling in biological entities. [16] These architectures might account for a greater percentage of quantum energy transfer, which occurs through electron transport and proton tunneling (usually in the form of hydrogen ions, H+). [17] [18] Tunneling refers to the ability of a subatomic particle to travel through potential energy barriers. [19] This ability is due, in part, to the principle of complementarity, which holds that certain substances have pairs of properties that cannot be measured separately without changing the outcome of measurement. Particles, such as electrons and protons, have wave-particle duality; they can pass through energy barriers due to their wave characteristics without violating the laws of physics. In order to quantify how quantum tunneling is used in many enzymatic activities, many biophysicists utilize the observation of hydrogen ions. When hydrogen ions are transferred, this is seen as a staple in an organelle's primary energy processing network; in other words, quantum effects are most usually at work in proton distribution sites at distances on the order of an angstrom (1 Å). [20] [21] In physics, a semiclassical (SC) approach is most useful in defining this process because of the transfer from quantum elements (e.g. particles) to macroscopic phenomena (e.g. biochemicals). Aside from hydrogen tunneling, studies also show that electron transfer between redox centers through quantum tunneling plays an important role in enzymatic activity of photosynthesis and cellular respiration (see also Mitochondria section below). [15] [18]
Ferritin
Ferritin is an iron storage protein that is found in plants and animals. It is usually formed from 24 subunits that self-assemble into a spherical shell that is approximately 2 nm thick, with an outer diameter that varies with iron loading up to about 16 nm. Up to ~4500 iron atoms can be stored inside the core of the shell in the Fe3+ oxidation state as water-insoluble compounds such as ferrihydrite and magnetite. [22] Ferritin is able to store electrons for at least several hours, which reduce the Fe3+ to water soluble Fe2+. [23] Electron tunneling as the mechanism by which electrons transit the 2 nm thick protein shell was proposed as early as 1988. [24] Electron tunneling and other quantum mechanical properties of ferritin were observed in 1992, [25] and electron tunneling at room temperature and ambient conditions was observed in 2005. [26] Electron tunneling associated with ferritin is a quantum biological process, and ferritin is a quantum biological agent.
Electron tunneling through ferritin between electrodes is independent of temperature, which indicates that it is substantially coherent and activation-less. [27] The electron tunneling distance is a function of the size of the ferritin. Single electron tunneling events can occur over distances of up to 8 nm through the ferritin, and sequential electron tunneling can occur up to 12 nm through the ferritin. It has been proposed that the electron tunneling is magnon-assisted and associated with magnetite microdomains in the ferritin core. [28]
Early evidence of quantum mechanical properties exhibited by ferritin in vivo was reported in 2004, where increased magnetic ordering of ferritin structures in placental macrophages was observed using small angle neutron scattering (SANS). [29] Quantum dot solids also show increased magnetic ordering in SANS testing, [30] and can conduct electrons over long distances. [31] Increased magnetic ordering of ferritin cores disposed in an ordered layer on a silicon substrate with SANS testing has also been observed. [32] Ferritin structures like those in placental macrophages have been tested in solid state configurations and exhibit quantum dot solid-like properties of conducting electrons over distances of up to 80 microns through sequential tunneling and formation of Coulomb blockades. [33] [34] [35] Electron transport through ferritin in placental macrophages may be associated with an anti-inflammatory function. [36]
Conductive atomic force microscopy of substantia nigra pars compacta (SNc) tissue demonstrated evidence of electron tunneling between ferritin cores, in structures that correlate to layers of ferritin outside of neuromelanin organelles. [37]
Evidence of ferritin layers in cell bodies of large dopamine neurons of the SNc and between those cell bodies in glial cells has also been found, [38] [39] [40] and is hypothesized to be associated with neuron function. [41] Overexpression of ferritin reduces the accumulation of reactive oxygen species, [42] and may act as a catalyst by increasing the ability of electrons from antioxidants to neutralize reactive oxygen species through electron tunneling. Ferritin has also been observed in ordered configurations in lysosomes associated with erythropoiesis, [43] where it may be associated with red blood cell production. While direct evidence of tunneling associated with ferritin in vivo in live cells has not yet been obtained, it may be possible to do so using quantum dots tagged with anti-ferritin, which should emit photons if electrons stored in the ferritin core tunnel to the quantum dots. [44]
 Electron microscope image of placental macrophage ferritin
Electron microscope image of placental macrophage ferritin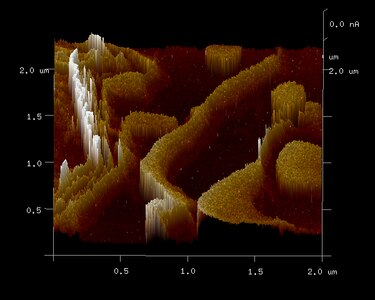 Conductive atomic force microscopy image of human substantia nigra pars compacta (SNc) tissue
Conductive atomic force microscopy image of human substantia nigra pars compacta (SNc) tissue Electron spectroscopic imaging of iron (red) outside of neuromelanin organelles
Electron spectroscopic imaging of iron (red) outside of neuromelanin organelles Electron microscope image of glial cell from SNc showing structures similar to ferritin in placental tissue
Electron microscope image of glial cell from SNc showing structures similar to ferritin in placental tissue








