
Endosomes are a collection of intracellular sorting organelles in eukaryotic cells. They are parts of endocytic membrane transport pathway originating from the trans Golgi network. Molecules or ligands internalized from the plasma membrane can follow this pathway all the way to lysosomes for degradation or can be recycled back to the cell membrane in the endocytic cycle. Molecules are also transported to endosomes from the trans Golgi network and either continue to lysosomes or recycle back to the Golgi apparatus.
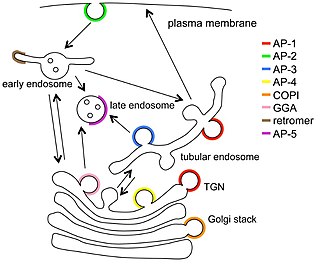
Vesicular transport adaptor proteins are proteins involved in forming complexes that function in the trafficking of molecules from one subcellular location to another. These complexes concentrate the correct cargo molecules in vesicles that bud or extrude off of one organelle and travel to another location, where the cargo is delivered. While some of the details of how these adaptor proteins achieve their trafficking specificity has been worked out, there is still much to be learned.
The coatomer is a protein complex that coats membrane-bound transport vesicles. Two types of coatomers are known:

Sortilin-related receptor, L(DLR class) A repeats containing is a protein that in humans is encoded by the SORL1 gene.

Sorting nexin-1 is a protein that in humans is encoded by the SNX1 gene. The protein encoded by this gene is a sorting nexin. SNX1 is a component of the retromer complex.

Sortilin (SORT1) is a protein that in humans is encoded by the SORT1 gene on chromosome 1. This protein is a type I membrane glycoprotein in the vacuolar protein sorting 10 protein (Vps10p) family of sorting receptors. While it is ubiquitously expressed in many tissues, sortilin is most abundant in the central nervous system. At the cellular level, sortilin functions in protein transport between the Golgi apparatus, endosome, lysosome, and plasma membrane, leading to its involvement in multiple biological processes such as glucose and lipid metabolism as well as neural development and cell death. Moreover, the function and role of sortilin is now emerging in several major human diseases such as hypertension, atherosclerosis, coronary artery disease, Alzheimer’s disease, and cancer. The SORT1 gene also contains one of 27 loci associated with increased risk of coronary artery disease.
BLOC-1 or biogenesis of lysosome-related organelles complex 1 is a ubiquitously expressed multisubunit protein complex in a group of complexes that also includes BLOC-2 and BLOC-3. BLOC-1 is required for normal biogenesis of specialized organelles of the endosomal-lysosomal system, such as melanosomes and platelet dense granules. These organelles are called LROs which are apparent in specific cell-types, such as melanocytes. The importance of BLOC-1 in membrane trafficking appears to extend beyond such LROs, as it has demonstrated roles in normal protein-sorting, normal membrane biogenesis, as well as vesicular trafficking. Thus, BLOC-1 is multi-purposed, with adaptable function depending on both organism and cell-type.

Syntaxin-6 is a protein that in humans is encoded by the STX6 gene.

Vesicle-associated membrane protein 3 is a protein that in humans is encoded by the VAMP3 gene.

Vacuolar protein sorting ortholog 35 (VPS35) is a protein involved in autophagy and is implicated in neurodegenerative diseases, such as Parkinson's disease (PD) and Alzheimer's disease (AD). VPS35 is part of a complex called the retromer, which is responsible for transporting select cargo proteins between vesicular structures and the Golgi apparatus. Mutations in the VPS35 gene (VPS35) cause aberrant autophagy, where cargo proteins fail to be transported and dysfunctional or unnecessary proteins fail to be degraded. There are numerous pathways affected by altered VPS35 levels and activity, which have clinical significance in neurodegeneration. There is therapeutic relevance for VPS35, as interventions aimed at correcting VPS35 function are in speculation.

Sorting nexin-2 is a protein that in humans is encoded by the SNX2 gene.

Vacuolar protein sorting-associated protein 26A is a protein that in humans is encoded by the VPS26A gene.

Rab11 family-interacting protein 5 is a protein that in humans is encoded by the RAB11FIP5 gene.
VPS29 is a human gene coding for the vacuolar protein sorting protein Vps29, a component of the retromer complex.

Sorting nexin-5 is a protein that in humans is encoded by the SNX5 gene.
Ras-related protein Rab-11B is a protein that in humans is encoded by the RAB11B gene. Rab11b is reported as most abundantly expressed in brain, heart and testes.
Sorting Nexin 6 also known as SNX6 is a well-conserved membrane-associated protein belonging to the sorting nexin family that is a component of the retromer complex. The protein contains a coiled-coil domain at its C terminus and a PX domain at its N terminus. Binding to PIM1 causes translocation to the nucleus. SNX6 has been shown to associate with TRAF4.

In the fields of biochemistry and cell biology, the cation-dependent mannose-6-phosphate receptor (CD-MPR) also known as the 46 kDa mannose 6-phosphate receptor is a protein that in humans is encoded by the M6PR gene.

Syntaxin-10 (STX10) is a SNARE protein that is encoded by the STX10 gene. This protein is found in most vertebrates but is noticeably absent from mice. As with other SNARE proteins, STX10 facilitates vesicle fusion and thus is important for intracellular trafficking of proteins and other cellular components. More specifically, STX10 has been implicated in endosome to Golgi trafficking of the mannose 6-phosphate receptor and glucose transporter type 4.

The SNX8 is a sorting nexin protein involved in intracellular molecular traffic from the early endosomes to the TGN. It is suggested that it acts as an adaptor protein in events related to immune response and cholesterol regulation, for example. As a protein of the SNXs family, the SNX8 is formed of 465 aminoacids and presents a BAR-domain and a PX-domain which are very relevant in relation to its functions. Furthermore, SNX8 study is motivated by its medical significance in relation to diseases such as Alzheimer's Disease, cancer, neurodevelopmental malformations and to its role in fighting against viral infections.


