| Common name | Binomial name | Order | Date of extinction | Former range | Picture |
|---|
| Broad-faced potoroo | Potorous platyops
Gould, 1844 | Diprotodontia | 1875 1 | Australia |  |
| Eastern hare wallaby | Lagorchestes leporides
Gould, 1841 | Diprotodontia | 1889 1 | 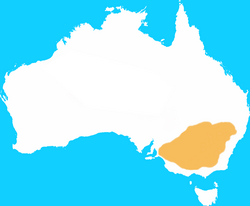
Australia |  |
| Lake Mackay hare-wallaby | Lagorchestes asomatus
Finlayson, 1943 | Diprotodontia | 1932 1 | Australia | |
| Desert rat-kangaroo | Caloprymnus campestris
Gould, 1843 | Diprotodontia | 1935 1 | 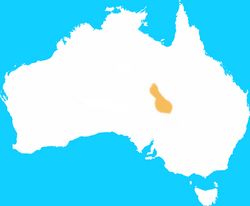
Australia |  |
Thylacine
or Tasmanian wolf/tiger | Thylacinus cynocephalus
Harris, 1808 | Dasyuromorphia | 1936 1 | 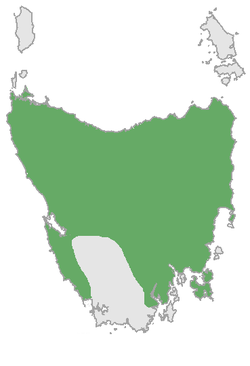
Australia, Tasmania |  |
| Toolache wallaby | Macropus greyi
Waterhouse, 1846 | Diprotodontia | 1939 1 | Australia |  |
| Desert bandicoot | Perameles eremiana
Spencer, 1837 | Peramelemorphia | 1943 1 | Australia |  |
| New South Wales barred bandicoot [16] | Perameles fasciata
Gray, 1841 | Peramelemorphia | mid-19th century | Australia |  |
| Southwestern barred bandicoot [16] | Perameles myosuros
Wagner, 1841 | Peramelemorphia | mid-19th century | Australia |  |
| Southern barred bandicoot [16] | Perameles notina
Thomas, 1922 | Peramelemorphia | mid-19th century | Australia | |
| Nullarbor barred bandicoot [16] | Perameles papillon
Travouillon & Phillips, 2018 | Peramelemorphia | early 20th century | Australia |  |
Lesser bilby
or yallara | Macrotis leucura
Thomas, 1887 | Peramelemorphia | 1960s 1 | 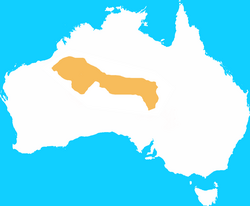
Australia |  |
| Southern pig-footed bandicoot | Chaeropus ecaudatus
Ogilby, 1838 | Peramelemorphia | 1950s 1 | 
Australia |  |
| Northern pig-footed bandicoot | Chaeropus yirratji
Travouillon et al., 2019 | Peramelemorphia | 1950s | |  |
| Crescent nail-tail wallaby | Onychogalea lunata
Gould, 1841 | Diprotodontia | 1956 1 | Australia (western and central) |  |
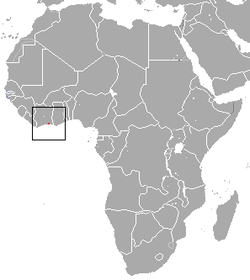
Red-bellied gracile opossum
or red-bellied gracile mouse opossum | Cryptonanus ignitus
Díaz, Flores and Barquez, 2002 | Didelphimorphia | 1962 1 | Argentina | |
| Nullarbor dwarf bettong | Bettongia pusilla
McNamara, 1997 | Diprotodontia | early 1500s 1 | Australia (Nullarbor Plain) | |
| Steller's sea cow | Hydrodamalis gigas
von Zimmermann, 1780 | Sirenia | 1768 1 | Commander Islands (Russia, United States) |  |
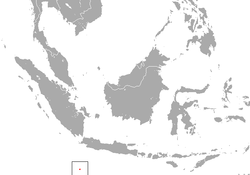
| Bramble Cay melomys | Melomys rubicola
Thomas, 1924 | Rodentia | 2016 1 | Australia (Bramble Cay) |  |
| Oriente cave rat | Boromys offella
Miller, 1916 | Rodentia | early 1500s 1 | Cuba |  |
| Torre's cave rat | Boromys torrei
Allen, 1917 | Rodentia | early 1500s 1 | Cuba |  |
| Imposter hutia | Hexolobodon phenax
Miller, 1929 | Rodentia | early 1500s 1 | Hispaniola (currently Haiti and the Dominican Republic) | |
| Montane hutia | Isolobodon montanus
Miller, 1922 | Rodentia | early 1500s 1 | Hispaniola | |
| Dwarf viscacha | Lagostomus crassus
Thomas, 1910 | Rodentia | early 1900s 1 | Peru | |
| Galápagos giant rat | Megaoryzomys curioi
Niethammer, 1964 | Rodentia | 1500s 1 | Santa Cruz Island (Galápagos) |  |
| Cuban coney | Geocapromys columbianus
Chapman, 1892 | Rodentia | early 1500s 1 | Cuba | |
| Hispaniolan edible rat | Brotomys voratus
Miller, 1916 | Rodentia | 1536–1546 1 | Hispaniola | |
| Puerto Rican hutia | Isolobodon portoricensis
Allen, 1916 | Rodentia | early 1900s 1 | Hispaniola; introduced to Puerto Rico, Saint Thomas Island, Saint Croix, U.S. Virgin Islands and Mona Island | |
| Big-eared hopping mouse | Notomys macrotis
Thomas, 1921 | Rodentia | 1843 1 | Australia (central Western Australia) | |
| Darling Downs hopping mouse | Notomys mordax
Thomas, 1921 | Rodentia | 1846 1 | Australia (Darling Downs, Queensland) | |
| White-footed rabbit-rat | Conilurus albipes
Lichtenstein, 1829 | Rodentia | early 1860s 1 | Australia (eastern coast) |  |
| Capricorn rabbit rat | Conilurus capricornensis
Cramb and Hocknull, 2010 | Rodentia | early 1500s 1 | Australia (Queensland) | |
| Short-tailed hopping mouse | Notomys amplus
Brazenor, 1936 | Rodentia | 1896 1 | Australia (Great Sandy Desert) | |
| Long-tailed hopping mouse | Notomys longicaudatus
Gould, 1844 | Rodentia | 1901 1 | Australia |  |
| Great hopping mouse | Notomys robustus
Mahoney, Smith and Medlin, 2008 | Rodentia | mid-1800s 1 | Australia (Flinders Ranges and Davenport Ranges) | |
Desmarest's pilorie
or Martinique giant rice rat | Megalomys desmarestii
Fischer, 1829 | Rodentia | 1902 1 | Martinique |  |
Saint Lucia pilorie
or Saint Lucia giant rice rat | Megalomys luciae
Major, 1901 | Rodentia | 1881 1 | Saint Lucia |  |
| Bulldog rat | Rattus nativitatis
Thomas, 1888 | Rodentia | 1903 1 | Christmas Island |  |
| Maclear's rat | Rattus macleari
Thomas, 1887 | Rodentia | 1903 1 | Christmas Island |  |
| Darwin's Galápagos mouse | Nesoryzomys darwini
Osgood, 1929 | Rodentia | 1930 1 | Galápagos Islands | |
| Gould's mouse | Pseudomys gouldii
Waterhouse, 1839 | Rodentia | 1930 1 | Australia (southern half) |  |
Plains rat
or palyoora | Pseudomys auritus
Thomas, 1910 | Rodentia | early 1800s 1 | Australia (Kangaroo Island and the Younghusband Peninsula) | |
| Pemberton's deer mouse | Peromyscus pembertoni
Burt, 1932 | Rodentia | 1931 1 | San Pedro Nolasco Island, Mexico |  |
| Samaná hutia | Plagiodontia ipnaeum
Johnson, 1948 | Rodentia | early 1500s [a] 1 | Hispaniola | |
| Hispaniola monkey | Antillothrix bernensis
MacPhee, Horovitz, Arredondo, & Jimenez Vasquez, 1995 | Primates | early 16th century | Hispaniola (currently Dominican Republic) | |
Lesser stick-nest rat
or white-tipped stick-nest rat | Leporillus apicalis
John Gould, 1854 | Rodentia | 1933 1 | Australia (west-central) |  |
| Indefatigable Galápagos mouse | Nesoryzomys indefessus
Thomas, 1899 | Rodentia | 1934 1 | Galápagos Islands | |
| Little Swan Island hutia | Geocapromys thoracatus
True, 1888 | Rodentia | 1955 1 | Swan Islands, Honduras |  |
| Blue-gray mouse | Pseudomys glaucus
Thomas, 1910 | Rodentia | 1956 1 | Australia (Queensland, New South Wales) | |
Buhler's coryphomys
or Buhler's rat | Coryphomys buehleri
Schaub, 1937 | Rodentia | early 1500s 1 | West Timor, Indonesia | |
| Insular cave rat | Heteropsomys insulans
Anthony, 1916 | Rodentia | early 1500s 1 | Vieques Island, Puerto Rico | |
| Candango mouse | Juscelinomys candango
Moojen, 1965 | Rodentia | 1960 1 | Central Brazil |  |
| Anthony's woodrat | Neotoma anthonyi
Allen, 1898 | Rodentia | 1926 1 | Isla Todos Santos, Mexico | |
| Bunker's woodrat | Neotoma bunkeri
Burt, 1932 | Rodentia | 1931 1 | Coronado Islands, Mexico | |
| Vespucci's rodent | Noronhomys vespuccii
Carleton and Olson, 1999 | Rodentia | 1500 1 | Fernando de Noronha, Brazil | |
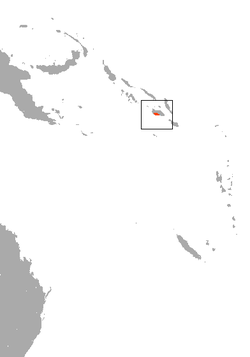
St. Vincent colilargo
or St. Vincent pygmy rice rat | Oligoryzomys victus
Thomas, 1898 | Rodentia | 1892 1 | Saint Vincent | |
| Jamaican rice rat | Oryzomys antillarum
Thomas, 1898 | Rodentia | 1877 1 | Jamaica |  |
| Nelson's rice rat | Oryzomys nelsoni
Merriam, 1889 | Rodentia | 1897 1 | Islas Marías, Mexico |  |
Nevis rice rat,
St. Eustatius rice rat, or St. Kitts rice rat | Pennatomys nivalis
Turvey, Weksler, Morris & Nokkert, 2010 | Rodentia | early 1500s [b] 1 | Sint Eustatius and Saint Kitts and Nevis |  |
| Christmas Island pipistrelle | Pipistrellus murrayi
Andrews, 1900 | Chiroptera | 2009 1 | Christmas Island | |
| Sardinian pika | Prolagus sardus
Wagner, 1832 | Lagomorpha | 1774 1 | Corsica and Sardinia |  |
| Marcano's solenodon | Solenodon marcanoi
Patterson, 1962 | Eulipotyphla | 1500s 1 | Dominican Republic | |
| Puerto Rican nesophontes | Nesophontes edithae
Anthony, 1916 | Eulipotyphla | early 1500s 1 | Puerto Rico, Vieques Island, Saint John, U.S. Virgin Islands and Saint Thomas, U.S. Virgin Islands |  |
| Atalaye nesophontes | Nesophontes hypomicrus
Miller, 1929 | Eulipotyphla | early 1500s 1 | Hispaniola | |
| Greater Cuban nesophontes | Nesophontes major
Arredondo, 1970 | Eulipotyphla | early 1500s 1 | Cuba | |
| Western Cuban nesophontes | Nesophontes micrus
Allen, 1917 | Eulipotyphla | early 1500s 1 | Cuba (including Isla de la Juventud) | |
| St. Michel nesophontes | Nesophontes paramicrus
Miller, 1929 | Eulipotyphla | early 1500s 1 | Hispaniola | |
| Haitian nesophontes | Nesophontes zamicrus
Miller, 1929 | Eulipotyphla | early 1500s 1 | Haiti | |
Lesser Mascarene flying fox
or dark flying fox | Pteropus subniger
kerr, 1792 | Chiroptera | 1864 1 | Réunion, Mauritius |  |
Guam flying fox
or Guam fruit bat | Pteropus tokudae
Tate, 1934 | Chiroptera | 1968 1 | Guam | |
Dusky flying fox
or Percy Island flying fox | Pteropus brunneus
Dobson, 1878 | Chiroptera | 1870 1 | Percy Islands (Australia) | |
| Large Palau flying fox | Pteropus pilosus
Andersen, 1908 | Chiroptera | 1874 1 | Palau | |
| Large sloth lemur | Palaeopropithecus ingens
Grandidier, 1899 | Primates | 1620 1 | 
In green |  |
| Aurochs | Bos primigenius
Bojanus, 1827 | Artiodactyla | 1627 1 |  |  |
| Bluebuck | Hippotragus leucophaeus
Pallas, 1766 | Artiodactyla | 1800 1 |  |  |
| Red gazelle | Eudorcas rufina
Thomas, 1894 | Artiodactyla | late 1800s 1 | Algeria |  |
| Schomburgk's deer | Rucervus schomburgki
Blyth, 1863 | Artiodactyla | 1932 1 | Thailand |  |
Queen of Sheba's gazelle
or Yemen gazelle | Gazella bilkis
Grover and Lay, 1985 | Artiodactyla | 1951 1 | Yemen | |
| Madagascan dwarf hippopotamus | Hippopotamus lemerlei
Milne-Edwards, 1868 | Artiodactyla | early 1500s [c] 1 | Madagascar |  |
| Falkland Islands wolf or warrah | Dusicyon australis
Kerr, 1792 | Carnivora | 1876 1 | Falkland Islands |  |
| Dusicyon avus | Dusicyon avus
Burmeister, 1866 | Carnivora | early 1500s 1 | Argentina, Chile, Brazil, Uruguay, Paraguay | |
| Sea mink | Neogale macrodon
Prentiss, 1903 | Carnivora | 1894 1 | United States (Maine, Massachusetts) and Canada (New Brunswick, Newfoundland) | |
| Japanese sea lion | Zalophus japonicus
Peters, 1866 | Carnivora | 1970s 1 | Japan, Korea, Russia |  |
| Caribbean monk seal | Neomonachus tropicalis
Gray, 1850 | Carnivora | 1952 1 | Caribbean Sea |  |
| Giant fossa | Cryptoprocta spelea
Grandidier, 1902 | Carnivora | before 1658 1 |  |  |
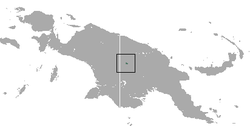
| Lord Howe long-eared bat | Nyctophilus howensis
McKean, 1975 | Chiroptera | prior to 1972 1 | Lord Howe Island, Australia |
| Japanese otter | Lutra nippon Imaizumi & Yoshiyuki, 1989 | Carnivora | 1990s [19] | Japan |  |