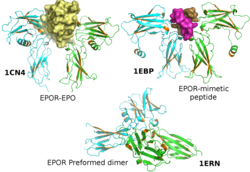
Erythropoietin, also known as erythropoetin, haematopoietin, or haemopoietin, is a glycoprotein cytokine secreted mainly by the kidneys in response to cellular hypoxia; it stimulates red blood cell production (erythropoiesis) in the bone marrow. Low levels of EPO are constantly secreted in sufficient quantities to compensate for normal red blood cell turnover. Common causes of cellular hypoxia resulting in elevated levels of EPO include any anemia, and hypoxemia due to chronic lung disease.

In oncology, polycythemia vera is an uncommon myeloproliferative neoplasm in which the bone marrow makes too many red blood cells.
The JAK-STAT signaling pathway is a chain of interactions between proteins in a cell, and is involved in processes such as immunity, cell division, cell death, and tumour formation. The pathway communicates information from chemical signals outside of a cell to the cell nucleus, resulting in the activation of genes through the process of transcription. There are three key parts of JAK-STAT signalling: Janus kinases (JAKs), signal transducer and activator of transcription proteins (STATs), and receptors. Disrupted JAK-STAT signalling may lead to a variety of diseases, such as skin conditions, cancers, and disorders affecting the immune system.

Tyrosine-protein phosphatase non-receptor type 11 (PTPN11) also known as protein-tyrosine phosphatase 1D (PTP-1D), Src homology region 2 domain-containing phosphatase-2 (SHP-2), or protein-tyrosine phosphatase 2C (PTP-2C) is an enzyme that in humans is encoded by the PTPN11 gene. PTPN11 is a protein tyrosine phosphatase (PTP) Shp2.

Signal transducer and activator of transcription 5 (STAT5) refers to two highly related proteins, STAT5A and STAT5B, which are part of the seven-membered STAT family of proteins. Though STAT5A and STAT5B are encoded by separate genes, the proteins are 90% identical at the amino acid level. STAT5 proteins are involved in cytosolic signalling and in mediating the expression of specific genes. Aberrant STAT5 activity has been shown to be closely connected to a wide range of human cancers, and silencing this aberrant activity is an area of active research in medicinal chemistry.
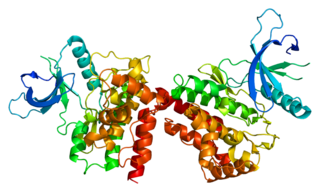
Janus kinase 2 is a non-receptor tyrosine kinase. It is a member of the Janus kinase family and has been implicated in signaling by members of the type II cytokine receptor family, the GM-CSF receptor family, the gp130 receptor family, and the single chain receptors.

The granulocyte-macrophage colony-stimulating factor receptor also known as CD116, is a receptor for granulocyte-macrophage colony-stimulating factor, which stimulates the production of white blood cells. In contrast to M-CSF and G-CSF which are lineage specific, GM-CSF and its receptor play a role in earlier stages of development. The receptor is primarily located on neutrophils, eosinophils and monocytes/macrophages, it is also on CD34+ progenitor cells (myeloblasts) and precursors for erythroid and megakaryocytic lineages, but only in the beginning of their development.
Tyrosine-protein kinase Lyn is a protein that in humans is encoded by the LYN gene.

Phosphatidylinositol 3-kinase regulatory subunit alpha is an enzyme that in humans is encoded by the PIK3R1 gene.
Tyrosine-protein phosphatase non-receptor type 6, also known as Src homology region 2 domain-containing phosphatase-1 (SHP-1), is an enzyme that in humans is encoded by the PTPN6 gene.
Suppressor of cytokine signaling 3 is a protein that in humans is encoded by the SOCS3 gene. This gene encodes a member of the STAT-induced STAT inhibitor (SSI), also known as suppressor of cytokine signaling (SOCS), family. SSI family members are cytokine-inducible negative regulators of cytokine signaling.

Crk-like protein is a protein that in humans is encoded by the CRKL gene.
Signal transducer and activator of transcription 5A is a protein that in humans is encoded by the STAT5A gene. STAT5A orthologs have been identified in several placentals for which complete genome data are available.

Src homology 2 (SH2) domain containing inositol polyphosphate 5-phosphatase 1(SHIP1) is an enzyme with phosphatase activity. SHIP1 is structured by multiple domain and is encoded by the INPP5D gene in humans. SHIP1 is expressed predominantly by hematopoietic cells but also, for example, by osteoblasts and endothelial cells. This phosphatase is important for the regulation of cellular activation. Not only catalytic but also adaptor activities of this protein are involved in this process. Its movement from the cytosol to the cytoplasmic membrane, where predominantly performs its function, is mediated by tyrosine phosphorylation of the intracellular chains of cell surface receptors that SHIP1 binds. Insufficient regulation of SHIP1 leads to different pathologies.

Tyrosine-protein kinase Tec is a tyrosine kinase that in humans is encoded by the TEC gene. Tec kinase is expressed in hematopoietic, liver, and kidney cells and plays an important role in T-helper cell processes. Tec kinase is the name-giving member of the Tec kinase family, a family of non-receptor protein-tyrosine kinases.

Macrophage-stimulating protein receptor is a protein that in humans is encoded by the MST1R gene. MST1R is also known as RON kinase, named after the French city in which it was discovered. It is related to the c-MET receptor tyrosine kinase.

SH2B adapter protein 3 (SH2B3), also known as lymphocyte adapter protein (LNK), is a protein that in humans is encoded by the SH2B3 gene on chromosome 12.

CFU-E stands for Colony Forming Unit-Erythroid. It arises from CFU-GEMM and gives rise to proerythroblasts.
Erythropoietin in neuroprotection is the use of the glycoprotein erythropoietin (Epo) for neuroprotection. Epo controls erythropoiesis, or red blood cell production.
Stephanie S. Watowich is an American immunologist. Watowich is the deputy chair of the Department of Immunology at MD Anderson Cancer Center in Houston, TX. She is a professor within the department as well and serves as the co-director of the Center for Inflammation and Cancer at the MD Anderson Cancer Center. Watowich’s research has focused on transcriptional control of innate immunity, with specific interest in the actions of the cytokine-activated STAT transcriptional regulators.