Testosterone is the primary male sex hormone and androgen in males.[4] In humans, testosterone plays a key role in the development of male reproductive tissues such as testicles and prostate, as well as promoting secondary sexual characteristics such as increased muscle and bone mass, and the growth of body hair. It is associated with increased aggression, sex drive, dominance, courtship display, and a wide range of behavioral characteristics.[5] In addition, testosterone in both sexes is involved in health and well-being, where it has a significant effect on overall mood, cognition, social and sexual behavior, metabolism and energy output, the cardiovascular system, and in the prevention of osteoporosis.[6][7] Insufficient levels of testosterone in men may lead to abnormalities including frailty, accumulation of adipose fat tissue within the body, anxiety and depression, sexual performance issues, and bone loss.
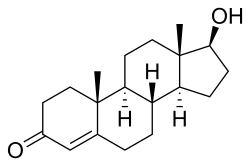
Testosterone is a steroid hormone from the androstane class containing a ketone and a hydroxyl group at positions three and seventeen respectively. It is biosynthesized in several steps from cholesterol and is converted in the liver to inactive metabolites.[9] It exerts its action through binding to and activation of the androgen receptor.[9] In humans and most other vertebrates, testosterone is secreted primarily by the testicles of males and, to a lesser extent, the ovaries of females. On average, in adult males, levels of testosterone are about seven to eight times as great as in adult females.[10] As the metabolism of testosterone in males is more pronounced, the daily production is about 20 times greater in men.[11][12] Females are also more sensitive to the hormone.[13][pageneeded]
Regarding aggression, testosterone does not create aggression in an individual, but amplifies aggressive behaviors and responses that are already learned by the individual, increasing sensitivity to social triggers rather than inventing violence.[17]
Biological effects
Effects on physiological development
In general, androgens such as testosterone promote protein synthesis and thus growth of tissues with androgen receptors.[18] Testosterone can be described as having anabolic and androgenic (virilising) effects, though these categorical descriptions are somewhat arbitrary, as there is a great deal of mutual overlap between them.[19] The relative potency of these effects can depend on various factors and is a topic of ongoing research.[20][21] Testosterone can either directly exert effects on target tissues or be metabolized by 5α-reductase into dihydrotestosterone (DHT) or aromatized to estradiol (E2).[20] Both testosterone and DHT bind to an androgen receptor; however, DHT has a stronger binding affinity than testosterone and may have more androgenic effect in certain tissues at lower levels.[20]
Testosterone effects can also be classified by the age of usual occurrence. For postnatal effects in both males and females, these are mostly dependent on the levels and duration of circulating free testosterone.[22]
Before birth
Effects before birth are divided into two categories, classified in relation to the stages of development.
During the second trimester, androgen level is associated with sex formation.[23] Specifically, testosterone, along with anti-Müllerian hormone (AMH) promote growth of the Wolffian duct and degeneration of the Müllerian duct respectively.[24] This period affects the femininization or masculinization of the fetus and can be a better predictor of feminine or masculine behaviours such as sex typed behaviour than an adult's own levels. Prenatal androgens apparently influence interests and engagement in gendered activities and have moderate effects on spatial abilities.[25] Among women with congenital adrenal hyperplasia, a male-typical play in childhood correlated with reduced satisfaction with the female gender and reduced heterosexual interest in adulthood.[26]
Early infancy
Early infancy androgen effects are the least understood. In the first weeks of life for male infants, testosterone levels rise. The levels remain in a pubertal range for a few months, but usually reach the barely detectable levels of childhood by 4–7 months of age.[27][28] The function of this rise in humans is unknown. It has been theorized that brain masculinization is occurring since no significant changes have been identified in other parts of the body.[29] The male brain is masculinized by the aromatization of testosterone into estradiol,[30] which crosses the blood–brain barrier and enters the male brain, whereas female fetuses have α-fetoprotein, which binds the estrogen so that female brains are not affected.[31]
Pubertal effects begin to occur when androgen has been higher than normal adult female levels for months or years. In males, these are usual late pubertal effects, and occur in women after prolonged periods of heightened levels of free testosterone in the blood. The effects include:[32][33]
Adult testosterone effects are more clearly demonstrable in males than in females, but are likely important to both sexes. Some of these effects may decline as testosterone levels might decrease in the later decades of adult life.[38]
The brain is also affected by this sexual differentiation;[23] the enzymearomatase converts testosterone into estradiol that is responsible for masculinization of the brain in male mice. In humans, masculinization of the fetal brain appears, by observation of gender preference in patients with congenital disorders of androgen formation or androgen receptor function, to be associated with functional androgen receptors.[39]
Testosterone does not appear to increase the risk of developing prostate cancer. In people who have undergone testosterone deprivation therapy, testosterone increases beyond the castrate level have been shown to increase the rate of spread of an existing prostate cancer.[41][42][43]
Conflicting results have been obtained concerning the importance of testosterone in maintaining cardiovascular health.[44][45] Nevertheless, maintaining normal testosterone levels in elderly men has been shown to improve many parameters that are thought to reduce cardiovascular disease risk, such as increased lean body mass, decreased visceral fat mass, decreased total cholesterol, and improved glycemic control.[46]
High androgen levels are associated with menstrual cycle irregularities in both clinical populations and healthy women.[bettersourceneeded][47] There also can be effects in unusual hair growth, acne, weight gain, infertility, and sometimes even scalp hair loss. These effects are seen largely in women with polycystic ovary syndrome (PCOS). For women with PCOS, hormones like birth control pills can be used to help lessen the effects of this increased level of testosterone.[48]
Attention, memory, and spatial ability are key cognitive functions affected by testosterone in humans. Preliminary evidence suggests that low testosterone levels may be a risk factor for cognitive decline and possibly for dementia of the Alzheimer's type,[49][50][51][52] a key argument in life extension medicine for the use of testosterone in anti-aging therapies. Much of the literature, however, suggests a curvilinear or even quadratic relationship between spatial performance and circulating testosterone,[53] where both hypo- and hypersecretion (deficient- and excessive-secretion) of circulating androgens have negative effects on cognition.
2020 guidelines from the American College of Physicians support the discussion of testosterone treatment in adult men with age-related low levels of testosterone who have sexual dysfunction. They recommend yearly evaluation regarding possible improvement and, if none, to discontinue testosterone; physicians should consider intramuscular treatments, rather than transdermal treatments, due to costs and since the effectiveness and harm of either method is similar. Testosterone treatment for reasons other than possible improvement of sexual dysfunction may not be recommended.[61][62] Current clinical guidelines recommend comprehensive baseline evaluation including complete blood count, lipid panel, prostate-specific antigen, and cardiovascular risk assessment before initiating testosterone replacement therapy.[63] Regular monitoring during treatment typically includes hematocrit levels every 3-6 months to prevent polycythemia, along with PSA monitoring in men over 40.[64][65]
Testosterone levels follow a circadian rhythm that peaks early each day, regardless of sexual activity.[66]
In women, correlations may exist between positive orgasm experience and testosterone levels. Studies have shown small or inconsistent correlations between testosterone levels and male orgasm experience, as well as sexual assertiveness in both sexes.[67][68]
Sexual arousal and masturbation in women produce small increases in testosterone concentrations.[69] The plasma levels of various steroids significantly increase after masturbation in men and the testosterone levels correlate to those levels.[70]
Mammalian studies
Studies conducted in rats have indicated that their degree of sexual arousal is sensitive to reductions in testosterone. When testosterone-deprived rats were given medium levels of testosterone, their sexual behaviours (copulation, partner preference, etc.) resumed, but not when given low amounts of the same hormone. Therefore, these mammals may provide a model for studying clinical populations among humans with sexual arousal deficits such as hypoactive sexual desire disorder.[71]
Every mammalian species examined demonstrated a marked increase in a male's testosterone level upon encountering a novel female. The reflexive testosterone increases in male mice is related to the male's initial level of sexual arousal.[72]
In non-human primates, it may be that testosterone in puberty stimulates sexual arousal, which allows the primate to increasingly seek out sexual experiences with females and thus creates a sexual preference for females.[73] Some research has also indicated that if testosterone is eliminated in an adult male human or other adult male primate's system, its sexual motivation decreases, but there is no corresponding decrease in ability to engage in sexual activity (mounting, ejaculating, etc.).[73]
In accordance with sperm competition theory, testosterone levels are shown to increase as a response to previously neutral stimuli when conditioned to become sexual in male rats.[74] This reaction engages penile reflexes (such as erection and ejaculation) that aid in sperm competition when more than one male is present in mating encounters, allowing for more production of successful sperm and a higher chance of reproduction.
Males
In men, higher levels of testosterone are associated with periods of sexual activity.[75][76]
Men who watch a sexually explicit movie have an average increase of 35% in testosterone, peaking at 60–90 minutes after the end of the film, but no increase is seen in men who watch sexually neutral films.[77] Men who watch sexually explicit films also report increased motivation and competitiveness, and decreased exhaustion.[78] A link has also been found between relaxation following sexual arousal and testosterone levels.[79]
Females
Androgens may modulate the physiology of vaginal tissue and contribute to female genital sexual arousal.[80] Women's level of testosterone is higher when measured pre-intercourse vs. pre-cuddling, as well as post-intercourse vs. post-cuddling.[81] There is a time lag effect when testosterone is administered, on genital arousal in women. In addition, a continuous increase in vaginal sexual arousal may result in higher genital sensations and sexual appetitive behaviors.[82]
Testosterone may prove to be an effective treatment in female sexual arousal disorders,[83] and is available as a dermal patch. There is no FDA-approved androgen preparation for the treatment of androgen insufficiency; however, it has been used as an off-label use to treat low libido and sexual dysfunction in older women. Testosterone may be a treatment for postmenopausal women as long as they are effectively estrogenized.[83]
Romantic relationships
Falling in love has been linked with decreases in men's testosterone levels while mixed changes are reported for women's testosterone levels.[84][85] There has been speculation that these changes in testosterone result in the temporary reduction of differences in behavior between the sexes.[85] However, the testosterone changes observed do not seem to be maintained as relationships develop over time.[84][85]
Men who produce less testosterone are more likely to be in a relationship[86] or married,[87] and men who produce more testosterone are more likely to divorce.[87] Marriage or commitment could cause a decrease in testosterone levels.[88] Single men who have not had relationship experience have lower testosterone levels than single men with experience. It is suggested that these single men with prior experience are in a more competitive state than their non-experienced counterparts.[89] Married men who engage in bond-maintenance activities such as spending the day with their spouse or child have no different testosterone levels compared to times when they do not engage in such activities. Collectively, these results suggest that the presence of competitive activities rather than bond-maintenance activities is more relevant to changes in testosterone levels.[90]
Men who produce more testosterone are more likely to engage in extramarital sex.[87] Testosterone levels do not rely on physical presence of a partner; testosterone levels of men engaging in same-city and long-distance relationships are similar.[86] Physical presence may be required for women who are in relationships for the testosterone–partner interaction, where same-city partnered women have lower testosterone levels than long-distance partnered women.[91]
Fatherhood
Fatherhood decreases testosterone levels in men, suggesting that the emotions and behaviour tied to paternal care decrease testosterone levels. In humans and other species that utilize allomaternal care, paternal investment in offspring is beneficial to said offspring's survival because it allows the two parents to raise multiple children simultaneously. This increases the reproductive fitness of the parents because their offspring are more likely to survive and reproduce. Paternal care increases offspring survival due to increased access to higher quality food and reduced physical and immunological threats.[92] This is particularly beneficial for humans since offspring are dependent on parents for extended periods of time and mothers have relatively short inter-birth intervals.[93]
While the extent of paternal care varies between cultures, higher investment in direct child care has been seen to be correlated with lower average testosterone levels as well as temporary fluctuations.[94] For instance, fluctuation in testosterone levels when a child is in distress has been found to be indicative of fathering styles. If a father's testosterone levels decrease in response to hearing their baby cry, it is an indication of empathizing with the baby. This is associated with increased nurturing behavior and better outcomes for the infant.[95]
Motivation
Testosterone levels play a major role in risk-taking during financial decisions.[96][97] Higher testosterone levels in men reduce the risk of becoming or staying unemployed.[98] Research has also found that heightened levels of testosterone and cortisol are associated with an increased risk of impulsive and violent criminal behavior.[99] On the other hand, elevated testosterone in men may increase their generosity, primarily to attract a potential mate.[100][101]
Most studies support a link between adult criminality and testosterone.[102][103][104][105] Nearly all studies of juvenile delinquency and testosterone are not significant. Most studies have found testosterone to be associated with behaviors or personality traits linked with antisocial behavior[106] and alcoholism. Many studies[which?] have been undertaken on the relationship between more general aggressive behavior, and feelings, and testosterone. About half of studies have found a relationship and about half, no relationship.[107] Studies have found that testosterone facilitates aggression by modulating vasopressin receptors in the hypothalamus.[108]
There are two theories on the role of testosterone in aggression and competition.[109] The first is the challenge hypothesis which states that testosterone would increase during puberty, thus facilitating reproductive and competitive behavior which would include aggression.[109] It is therefore the challenge of competition among males that facilitates aggression and violence.[109] Studies conducted have found direct correlation between testosterone and dominance, especially among the most violent criminals in prison who had the highest testosterone.[109] The same research found fathers (outside competitive environments) had the lowest testosterone levels compared to other males.[109]
The second theory is similar and known as "evolutionary neuroandrogenic (ENA) theory of male aggression".[110][111] Testosterone and other androgens have evolved to motivate men to pursue competition, even when doing so leads to risk. By doing so, individuals with masculinized brains are better able to survive and copulate with as many mates as possible.[110] The masculinization of the brain is not just mediated by testosterone levels at the adult stage, but also testosterone exposure in the womb. Higher pre-natal testosterone indicated by a low digit ratio as well as adult testosterone levels increased risk of fouls or aggression among male players in a soccer game.[112] Studies have found higher pre-natal testosterone or lower digit ratio to be correlated with higher aggression.[113][114][115][116][117]
The rise in testosterone during competition predicted aggression in males, but not in females.[118] In one experiment, subjects who interacted with handguns showed higher testosterone levels and aggression than those who interacted with toys.[119] One study proposed that natural selection may have caused men to be more sensitive to situations in which their status is challenged, and that testosterone is the key factor that causes these situations to spark into aggression.[120] Structural brain characteristics associated with testosterone levels have also been found to predict individuals' levels of aggression.[121]
The Annals of the New York Academy of Sciences has found that the use of anabolic steroids (which increases testosterone) among teenagers is correlated with increased likelihood of using violence.[122] One study found that administering testosterone increased verbal aggression in some participants.[123]
A few studies indicate that the testosterone derivative estradiol might play an important role in male aggression.[107][124][125][126] Estradiol is known to correlate with aggression in male mice.[127] Moreover, the conversion of testosterone to estradiol regulates male aggression in sparrows during breeding season.[128] Rats who were given anabolic steroids that increase testosterone were also more physically aggressive to provocation as a result of "threat sensitivity".[129]
The relationship between testosterone and aggression may also function indirectly, as it has been proposed that testosterone does not amplify tendencies towards aggression, but rather amplifies whatever tendencies will allow an individual to maintain social status when challenged. In most animals, aggression is the means of maintaining social status. However, humans have multiple ways of obtaining status. This could explain why some studies find a link between testosterone and pro-social behaviour, if pro-social behaviour is rewarded with social status. Thus the link between testosterone and aggression and violence is due to these being rewarded with social status.[130] The relationship may also be one of a "permissive effect" whereby testosterone does elevate aggression levels, but only in the sense of allowing average aggression levels to be maintained; chemically or physically castrating the individual will reduce aggression levels (though not eliminate them) but the individual only needs a small-level of pre-castration testosterone to have aggression levels to return to normal, which they will remain at even if additional testosterone is added. Testosterone may also simply exaggerate or amplify existing aggression; for example, chimpanzees who receive testosterone increases become more aggressive to chimps lower than them in the social hierarchy, but will still be submissive to chimps higher than them. Testosterone thus does not make the chimpanzee indiscriminately aggressive, but instead amplifies his pre-existing aggression towards lower-ranked chimps.[131]
In humans, testosterone appears more to promote status-seeking and social dominance than simply increasing physical aggression. When controlling for the effects of belief in having received testosterone, women who have received testosterone make fairer offers than women who have not received testosterone.[132]
Robert Sapolsky explains by citing different studies that, testosterone does not "invent" aggression. Instead, it acts as an amplifier, increasing the "volume" of an individual's sensitivity to social triggers for aggression that they have already learned. Aggression can persist even after testosterone is removed (for example, by castration) if someone has prior experience with aggressive behavior, showing hormones are not the sole cause. Studies in animals show testosterone reinforces existing social hierarchies rather than overturning them, making individuals more aggressive only toward those already below them, but not challenging those who are above them in hierarchy. According to the Challenge Hypothesis, testosterone mainly motivates behavior that helps maintain status when it is threatened. In humans, this can lead to pro-social outcomes—such as generosity, honesty, or cooperation—when those traits bring respect and help them maintain their status in society (as status in human society does not come only through aggression, but through other means too), meaning testosterone’s effects depend heavily on social context. Ultimately societal norms, not testosterone itself, determine whether aggression or cooperation is rewarded.[17]
Fairness
Testosterone might encourage fair behavior. For one study, subjects took part in a behavioral experiment where the distribution of a real amount of money was decided. The rules allowed both fair and unfair offers. The negotiating partner could subsequently accept or decline the offer. The fairer the offer, the less probable a refusal by the negotiating partner. If no agreement was reached, neither party earned anything. Test subjects with an artificially enhanced testosterone level generally made better, fairer offers than those who received placebos, thus reducing the risk of a rejection of their offer to a minimum. Two later studies have empirically confirmed these results.[133][134][135] However men with high testosterone were significantly 27% less generous in an ultimatum game.[136] This additional information could suggest, contrarily, that testosterone may encourage greed or selfishness. Fairer offers from test subjects with higher testosterone in the original study increase the likeliness of the offer being accepted by the negotiating partner, therefore decreasing the probability of both participants leaving without any money.
Biological activity
Free testosterone
Lipophilichormones (soluble in lipids but not in water), such as steroid hormones, including testosterone, are transported in water-based blood plasma through specific and non-specific proteins. Specific proteins include sex hormone-binding globulin (SHBG), which binds testosterone, dihydrotestosterone, estradiol, and other sex steroids. Non-specific binding proteins include albumin. The part of the total hormone concentration that is not bound to its respective specific carrier protein is the free part. As a result, testosterone which is not bound to SHBG is called free testosterone. Only the free amount of testosterone can bind to an androgenic receptor, which means it has biological activity.[137] While a significant portion of testosterone is bound to SHBG, a small fraction of testosterone (1%-2%)[138] is bound to albumin and the binding of testosterone to albumin is weak and can be reversed easily;[139][140] as such, both albumin-bound and unbound testosterone are considered to be bioavailable testosterone.[139][140] This binding plays an important role in regulating the transport, tissue delivery, bioactivity, and metabolism of testosterone.[140][139] At the tissue level, testosterone dissociates from albumin and quickly diffuses into the tissues. The percentage of testosterone bound to SHBG is lower in men than in women. Both the free fraction and the one bound to albumin are available at the tissue level (their sum constitutes the bioavailable testosterone), while SHBG effectively and irreversibly inhibits the action of testosterone.[138] The relationship between sex steroids and SHBG in physiological and pathological conditions is complex, as various factors may influence the levels of plasma SHBG, affecting bioavailability of testosterone.[141][142][143]
Free testosterone (T) is transported into the cytoplasm of target tissuecells, where it can bind to the androgen receptor, or can be reduced to 5α-dihydrotestosterone (5α-DHT) by the cytoplasmic enzyme 5α-reductase. 5α-DHT binds to the same androgen receptor even more strongly than testosterone, so that its androgenic potency is about 5 times that of T.[149] The T-receptor or DHT-receptor complex undergoes a structural change that allows it to move into the cell nucleus and bind directly to specific nucleotide sequences of the chromosomal DNA. The areas of binding are called hormone response elements (HREs), and influence transcriptional activity of certain genes, producing the androgen effects.
Androgen receptors occur in many different vertebrate body system tissues, and both males and females respond similarly to similar levels. Greatly differing amounts of testosterone prenatally, at puberty, and throughout life account for a share of biological differences between males and females.
The bones and the brain are two important tissues in humans where the primary effect of testosterone is by way of aromatization to estradiol. In the bones, estradiol accelerates ossification of cartilage into bone, leading to closure of the epiphyses and conclusion of growth. In the central nervous system, testosterone is aromatized to estradiol. Estradiol rather than testosterone serves as the most important feedback signal to the hypothalamus (especially affecting LH secretion).[150][failed verification] In many mammals, prenatal or perinatal "masculinization" of the sexually dimorphic areas of the brain by estradiol derived from testosterone programs later male sexual behavior.[151]
Testosterone is an antagonist of the sigma-1 receptor (Ki = 1,014 or 201nM).[156] However, the concentrations of testosterone required for binding the receptor are far above even total circulating concentrations of testosterone in adult males (which range between 10 and 35nM).[157]
Biochemistry
Figure 1: Human steroidogenesis, showing testosterone near bottom
The largest amounts of testosterone (>95%) are produced by the testes in men,[5] while the adrenal glands account for most of the remainder. Testosterone is also synthesized in far smaller total quantities in women by the adrenal glands, thecal cells of the ovaries, and, during pregnancy, by the placenta.[160] In the testes, testosterone is produced by the Leydig cells.[161] The male generative glands also contain Sertoli cells, which require testosterone for spermatogenesis. Like most hormones, testosterone is supplied to target tissues in the blood where much of it is transported bound to a specific plasma protein, sex hormone-binding globulin (SHBG).
Notes: "The concentration of a steroid in the circulation is determined by the rate at which it is secreted from glands, the rate of metabolism of precursor or prehormones into the steroid, and the rate at which it is extracted by tissues and metabolized. The secretion rate of a steroid refers to the total secretion of the compound from a gland per unit time. Secretion rates have been assessed by sampling the venous effluent from a gland over time and subtracting out the arterial and peripheral venous hormone concentration. The metabolic clearance rate of a steroid is defined as the volume of blood that has been completely cleared of the hormone per unit time. The production rate of a steroid hormone refers to entry into the blood of the compound from all possible sources, including secretion from glands and conversion of prohormones into the steroid of interest. At steady state, the amount of hormone entering the blood from all sources will be equal to the rate at which it is being cleared (metabolic clearance rate) multiplied by blood concentration (production rate = metabolic clearance rate × concentration). If there is little contribution of prohormone metabolism to the circulating pool of steroid, then the production rate will approximate the secretion rate." Sources: See template.
The amount of testosterone synthesized is regulated by the hypothalamic–pituitary–testicular axis(Figure 2).[163] When testosterone levels are low, gonadotropin-releasing hormone (GnRH) is released by the hypothalamus, which in turn stimulates the pituitary gland to release FSH and LH. These latter two hormones stimulate the testis to synthesize testosterone. Finally, increasing levels of testosterone through a negative feedback loop act on the hypothalamus and pituitary to inhibit the release of GnRH and FSH/LH, respectively.
Factors affecting testosterone levels may include:
Exercise: Resistance training increases testosterone levels acutely,[167] however, in older men, that increase can be avoided by protein ingestion.[168]Endurance training in men may lead to lower testosterone levels.[169]
Weight loss: Reduction in weight may result in an increase in testosterone levels. Fat cells synthesize the enzyme aromatase, which converts testosterone, the male sex hormone, into estradiol, the female sex hormone.[175] However no clear association between body mass index and testosterone levels has been found.[176]
Behavior: Dominance challenges can, in some cases, stimulate increased testosterone release in men.[178]
Foods: Natural or man-made antiandrogens including spearmint tea reduce testosterone levels.[179][180][181]Licorice can decrease the production of testosterone and this effect is greater in females.[182]
In the hepatic 17-ketosteroid pathway of testosterone metabolism, testosterone is converted in the liver by 5α-reductase and 5β-reductase into 5α-DHT and the inactive 5β-DHT, respectively.[2][185] Then, 5α-DHT and 5β-DHT are converted by 3α-HSD into 3α-androstanediol and 3α-etiocholanediol, respectively.[2][185] Subsequently, 3α-androstanediol and 3α-etiocholanediol are converted by 17β-HSD into androsterone and etiocholanolone, which is followed by their conjugation and excretion.[2][185]3β-Androstanediol and 3β-etiocholanediol can also be formed in this pathway when 5α-DHT and 5β-DHT are acted upon by 3β-HSD instead of 3α-HSD, respectively, and they can then be transformed into epiandrosterone and epietiocholanolone, respectively.[187][188] A small portion of approximately 3% of testosterone is reversibly converted in the liver into androstenedione by 17β-HSD.[186]
In addition to conjugation and the 17-ketosteroid pathway, testosterone can also be hydroxylated and oxidized in the liver by cytochrome P450enzymes, including CYP3A4, CYP3A5, CYP2C9, CYP2C19, and CYP2D6.[189] 6β-Hydroxylation and to a lesser extent 16β-hydroxylation are the major transformations.[189] The 6β-hydroxylation of testosterone is catalyzed mainly by CYP3A4 and to a lesser extent CYP3A5 and is responsible for 75 to 80% of cytochrome P450-mediated testosterone metabolism.[189] In addition to 6β- and 16β-hydroxytestosterone, 1β-, 2α/β-, 11β-, and 15β-hydroxytestosterone are also formed as minor metabolites.[189][190] Certain cytochrome P450 enzymes such as CYP2C9 and CYP2C19 can also oxidize testosterone at the C17 position to form androstenedione.[189]
Two of the immediate metabolites of testosterone, 5α-DHT and estradiol, are biologically important and can be formed both in the liver and in extrahepatic tissues.[185] Approximately 5 to 7% of testosterone is converted by 5α-reductase into 5α-DHT, with circulating levels of 5α-DHT about 10% of those of testosterone, and approximately 0.3% of testosterone is converted into estradiol by aromatase.[5][185][191][192] 5α-Reductase is highly expressed in the male reproductive organs (including the prostate gland, seminal vesicles, and epididymides),[193]skin, hair follicles, and brain[194] and aromatase is highly expressed in adipose tissue, bone, and the brain.[195][196] As much as 90% of testosterone is converted into 5α-DHT in so-called androgenic tissues with high 5α-reductase expression,[186] and due to the several-fold greater potency of 5α-DHT as an AR agonist relative to testosterone,[197] it has been estimated that the effects of testosterone are potentiated 2- to 3-fold in such tissues.[198]
Levels
Total levels of testosterone in the body have been reported as 264 to 916ng/dL (nanograms per deciliter) in non-obese European and American men age 19 to 39years,[199] while mean testosterone levels in adult men have been reported as 630ng/dL.[200] Although commonly used as a reference range,[201] some physicians have disputed the use of this range to determine hypogonadism.[202][203] Several professional medical groups have recommended that 350ng/dL generally be considered the minimum normal level,[204] which is consistent with previous findings.[205][non-primary source needed][medical citation needed] Levels of testosterone in men decline with age.[199] In women, mean levels of total testosterone have been reported to be 32.6ng/dL.[206][207] In women with hyperandrogenism, mean levels of total testosterone have been reported to be 62.1ng/dL.[206][207]
In measurements of testosterone in blood samples, different assay techniques can yield different results.[212][213] Immunofluorescence assays exhibit considerable variability in quantifying testosterone concentrations in blood samples due to the cross-reaction of structurally similar steroids, leading to overestimating the results. In contrast, the liquid chromatography/tandem mass spectrometry method is more desirable: it offers superior specificity and precision, making it a more suitable choice for this application.[214]
Testosterone's bioavailable concentration is commonly determined using the Vermeulen calculation or more precisely using the modified Vermeulen method,[215][216] which considers the dimeric form of sex hormone-binding globulin.[217]
Both methods use chemical equilibrium to derive the concentration of bioavailable testosterone: in circulation, testosterone has two major binding partners, albumin (weakly bound) and sex hormone-binding globulin (strongly bound). These methods are described in detail in the accompanying figure.
Dimeric sex hormone-binding globulin with its testosterone ligands
Two methods for determining the concentration of bioavailable testosterone
Distribution
Testosterone has been detected at variably higher and lower levels among men of various nations and from various backgrounds, explanations for the causes of this have been relatively diverse.[218][219]
History and production
Nobel Prize winner, Leopold Ružička of Ciba, a pharmaceutical industry giant that synthesized testosterone
A testicular action was linked to circulating blood fractions– now understood to be a family of androgenic hormones– in the early work on castration and testicular transplantation in fowl by Arnold Adolph Berthold (1803–1861).[220] Research on the action of testosterone received a brief boost in 1889, when the Harvard professor Charles-Édouard Brown-Séquard (1817–1894), then in Paris, self-injected subcutaneously a "rejuvenating elixir" consisting of an extract of dog and guinea pig testicle. He reported in The Lancet that his vigor and feeling of well-being were markedly restored but the effects were transient,[221] and Brown-Séquard's hopes for the compound were dashed. Suffering the ridicule of his colleagues, he abandoned his work on the mechanisms and effects of androgens in human beings.
In 1927, the University of Chicago's Professor of Physiologic Chemistry, Fred C. Koch, established easy access to a large source of bovine testicles– the Chicago stockyards– and recruited students willing to endure the tedious work of extracting their isolates. In that year, Koch and his student, Lemuel McGee, derived 20mg of a substance from a supply of 40 pounds of bovine testicles that, when administered to castrated roosters, pigs and rats, re-masculinized them.[222] The group of Ernst Laqueur at the University of Amsterdam purified testosterone from bovine testicles in a similar manner in 1934, but the isolation of the hormone from animal tissues in amounts permitting serious study in humans was not feasible until three European pharmaceutical giants– Schering (Berlin, Germany), Organon (Oss, Netherlands) and Ciba– began full-scale steroid research and development programs in the 1930s.
The Organon group in the Netherlands were the first to isolate the hormone, identified in a May 1935 paper "On Crystalline Male Hormone from Testicles (Testosterone)".[223] They named the hormone testosterone, from the stems of testicle and sterol, and the suffix of ketone. The structure was worked out by Schering's Adolf Butenandt, at the Chemisches Institut of Technical University in Gdańsk.[224][225]
The chemical synthesis of testosterone from cholesterol was achieved in August that year by Butenandt and Hanisch.[226] Only a week later, the Ciba group in Zurich, Leopold Ruzicka (1887–1976) and A. Wettstein, published their synthesis of testosterone.[227] These independent partial syntheses of testosterone from a cholesterol base earned both Butenandt and Ruzicka the joint 1939 Nobel Prize in Chemistry.[225][228] Testosterone was identified as 17β-hydroxyandrost-4-en-3-one (C19H28O2), a solid polycyclic alcohol with a hydroxyl group at the 17th carbon atom. This also made it obvious that additional modifications on the synthesized testosterone could be made, i.e., esterification and alkylation.
The partial synthesis in the 1930s of abundant, potent testosterone esters permitted the characterization of the hormone's effects, so that Kochakian and Murlin (1936) were able to show that testosterone raised nitrogen retention (a mechanism central to anabolism) in the dog, after which Allan Kenyon's group[229] was able to demonstrate both anabolic and androgenic effects of testosterone propionate in eunuchoidal men, boys, and women. The period of the early 1930s to the 1950s has been called "The Golden Age of Steroid Chemistry",[230] and work during this period progressed quickly.[231]
Like other androsteroids, testosterone is manufactured industrially from microbial fermentation of plant cholesterol (e.g., from soybean oil). In the early 2000s, the steroid market weighed around one million tonnes and was worth $10 billion, making it the 2nd largest biopharmaceutical market behind antibiotics.[232]
Other species
Testosterone is observed in most vertebrates. Testosterone and the classical nuclear androgen receptor first appeared in gnathostomes (jawed vertebrates).[233]Agnathans (jawless vertebrates) such as lampreys do not produce testosterone but instead use androstenedione as a male sex hormone.[234]Fish make a slightly different form called 11-ketotestosterone.[235] Its counterpart in insects is ecdysone.[236] The presence of these ubiquitous steroids in a wide range of animals suggest that sex hormones have an ancient evolutionary history.[237]
↑Tuck SP, Francis RM (2009). "Testosterone, bone and osteoporosis". Advances in the Management of Testosterone Deficiency. Frontiers of Hormone Research. Vol.37. pp.123–32. doi:10.1159/000176049. ISBN978-3-8055-8622-1. PMID19011293.
↑Southren AL, Gordon GG, Tochimoto S, Pinzon G, Lane DR, Stypulkowski W (May 1967). "Mean plasma concentration, metabolic clearance and basal plasma production rates of testosterone in normal young men and women using a constant infusion procedure: effect of time of day and plasma concentration on the metabolic clearance rate of testosterone". The Journal of Clinical Endocrinology & Metabolism. 27 (5): 686–94. doi:10.1210/jcem-27-5-686. PMID6025472.
↑Southren AL, Tochimoto S, Carmody NC, Isurugi K (November 1965). "Plasma production rates of testosterone in normal adult men and women and in patients with the syndrome of feminizing testes". The Journal of Clinical Endocrinology & Metabolism. 25 (11): 1441–50. doi:10.1210/jcem-25-11-1441. PMID5843701.
123456789"Testosterone". Drugs.com. American Society of Health-System Pharmacists. December 4, 2015. Archived from the original on August 20, 2016. Retrieved September 3, 2016.
↑Liverman CT, Blazer DG, etal. (Institute of Medicine (US) Committee on Assessing the Need for Clinical Trials of Testosterone Replacement Therapy) (2004). "Introduction". Testosterone and Aging: Clinical Research Directions (Report). National Academies Press (US). Archived from the original on January 10, 2016. Retrieved September 26, 2016.
12Swaab DF, Garcia-Falgueras A (2009). "Sexual differentiation of the human brain in relation to gender identity and sexual orientation". Functional Neurology. 24 (1): 17–28. PMID19403051.
↑Hines M, Brook C, Conway GS (February 2004). "Androgen and psychosexual development: core gender identity, sexual orientation and recalled childhood gender role behavior in women and men with congenital adrenal hyperplasia (CAH)". Journal of Sex Research. 41 (1): 75–81. doi:10.1080/00224490409552215. PMID15216426. S2CID33519930.
↑Forest MG, Cathiard AM, Bertrand JA (July 1973). "Evidence of testicular activity in early infancy". The Journal of Clinical Endocrinology & Metabolism. 37 (1): 148–51. doi:10.1210/jcem-37-1-148. PMID4715291.
↑Corbier P, Edwards DA, Roffi J (1992). "The neonatal testosterone surge: a comparative study". Archives Internationales de Physiologie, de Biochimie et de Biophysique. 100 (2): 127–31. doi:10.3109/13813459209035274. PMID1379488.
↑Dakin CL, Wilson CA, Kalló I, Coen CW, Davies DC (May 2008). "Neonatal stimulation of 5-HT(2) receptors reduces androgen receptor expression in the rat anteroventral periventricular nucleus and sexually dimorphic preoptic area". The European Journal of Neuroscience. 27 (9): 2473–80. doi:10.1111/j.1460-9568.2008.06216.x. PMID18445234. S2CID23978105.
↑Kalat JW (2009). "Reproductive behaviors". Biological psychology. Belmont, Calif: Wadsworth, Cengage Learning. p.321. ISBN978-0-495-60300-9. Archived from the original on January 11, 2023. Retrieved October 8, 2020.
↑Morgentaler A, Schulman C (2009). "Testosterone and prostate safety". Advances in the Management of Testosterone Deficiency. Frontiers of Hormone Research. Vol.37. pp.197–203. doi:10.1159/000176054. ISBN978-3-8055-8622-1. PMID19011298.
↑Rhoden EL, Averbeck MA, Teloken PE (September 2008). "Androgen replacement in men undergoing treatment for prostate cancer". The Journal of Sexual Medicine. 5 (9): 2202–08. doi:10.1111/j.1743-6109.2008.00925.x. PMID18638000.
↑Morgentaler A, Traish AM (February 2009). "Shifting the paradigm of testosterone and prostate cancer: the saturation model and the limits of androgen-dependent growth". European Urology. 55 (2): 310–20. doi:10.1016/j.eururo.2008.09.024. PMID18838208.
↑Haddad RM, Kennedy CC, Caples SM, Tracz MJ, Boloña ER, Sideras K, etal. (January 2007). "Testosterone and cardiovascular risk in men: a systematic review and meta-analysis of randomized placebo-controlled trials". Mayo Clinic Proceedings. 82 (1): 29–39. doi:10.4065/82.1.29. PMID17285783.
↑Jones TH, Saad F (December 2009). "The effects of testosterone on risk factors for, and the mediators of, the atherosclerotic process". Atherosclerosis. 207 (2): 318–27. doi:10.1016/j.atherosclerosis.2009.04.016. PMID19464009.
↑Rosario ER, Chang L, Stanczyk FZ, Pike CJ (September 2004). "Age-related testosterone depletion and the development of Alzheimer disease". JAMA. 292 (12): 1431–32. doi:10.1001/jama.292.12.1431-b. PMID15383512.
↑Hogervorst E, Bandelow S, Combrinck M, Smith AD (2004). "Low free testosterone is an independent risk factor for Alzheimer's disease". Experimental Gerontology. 39 (11–12): 1633–39. doi:10.1016/j.exger.2004.06.019. PMID15582279. S2CID24803152.
↑Moffat SD, Hampson E (April 1996). "A curvilinear relationship between testosterone and spatial cognition in humans: possible influence of hand preference". Psychoneuroendocrinology. 21 (3): 323–37. doi:10.1016/0306-4530(95)00051-8. PMID8817730. S2CID7135870.
↑Lincoff AM, Bhasin S, Flevaris P, Mitchell LM, Basaria S, Boden WE, etal. (July 2023). "Cardiovascular Safety of Testosterone-Replacement Therapy". The New England Journal of Medicine. 389 (2): 107–117. doi:10.1056/NEJMoa2215025. PMID37326322. S2CID259176370.
↑Qaseem A, Horwitch CA, Vijan S, Etxeandia-Ikobaltzeta I, Kansagara D (January 2020). "Testosterone Treatment in Adult Men With Age-Related Low Testosterone: A Clinical Guideline From the American College of Physicians". Annals of Internal Medicine. 172 (2): 126–133. doi:10.7326/M19-0882. PMID31905405.
↑Bhasin S, Brito JP, Cunningham GR, Hayes FJ, Hodis HN, Matsumoto AM, etal. (May 2018). "Testosterone Therapy in Men With Hypogonadism: An Endocrine Society Clinical Practice Guideline". The Journal of Clinical Endocrinology and Metabolism. 103 (5): 1715–1744. doi:10.1210/jc.2018-00229. PMID29562364.
↑Mulhall JP, Trost LW, Brannigan RE, Kurtz EG, Redmon JB, Chiles KA, etal. (August 2018). "Evaluation and Management of Testosterone Deficiency: AUA Guideline". The Journal of Urology. 200 (2): 423–432. doi:10.1016/j.juro.2018.03.115. PMID29601923.
↑Fox CA, Ismail AA, Love DN, Kirkham KE, Loraine JA (January 1972). "Studies on the relationship between plasma testosterone levels and human sexual activity". The Journal of Endocrinology. 52 (1): 51–8. doi:10.1677/joe.0.0520051. PMID5061159.
↑Exton MS, Bindert A, Krüger T, Scheller F, Hartmann U, Schedlowski M (1999). "Cardiovascular and endocrine alterations after masturbation-induced orgasm in women". Psychosomatic Medicine. 61 (3): 280–89. doi:10.1097/00006842-199905000-00005. PMID10367606.
↑Purvis K, Landgren BM, Cekan Z, Diczfalusy E (September 1976). "Endocrine effects of masturbation in men". The Journal of Endocrinology. 70 (3): 439–44. doi:10.1677/joe.0.0700439. PMID135817.
↑Harding SM, Velotta JP (May 2011). "Comparing the relative amount of testosterone required to restore sexual arousal, motivation, and performance in male rats". Hormones and Behavior. 59 (5): 666–73. doi:10.1016/j.yhbeh.2010.09.009. PMID20920505. S2CID1577450.
↑James PJ, Nyby JG, Saviolakis GA (September 2006). "Sexually stimulated testosterone release in male mice (Mus musculus): roles of genotype and sexual arousal". Hormones and Behavior. 50 (3): 424–31. doi:10.1016/j.yhbeh.2006.05.004. PMID16828762. S2CID36436418.
↑Hart BL (December 1983). "Role of testosterone secretion and penile reflexes in sexual behavior and sperm competition in male rats: a theoretical contribution". Physiology & Behavior. 31 (6): 823–27. doi:10.1016/0031-9384(83)90279-2. PMID6665072. S2CID42155431.
↑Kraemer HC, Becker HB, Brodie HK, Doering CH, Moos RH, Hamburg DA (March 1976). "Orgasmic frequency and plasma testosterone levels in normal human males". Archives of Sexual Behavior. 5 (2): 125–32. doi:10.1007/BF01541869. PMID1275688. S2CID38283107.
↑Pirke KM, Kockott G, Dittmar F (November 1974). "Psychosexual stimulation and plasma testosterone in man". Archives of Sexual Behavior. 3 (6): 577–84. doi:10.1007/BF01541140. PMID4429441. S2CID43495791.
↑Tuiten A, Van Honk J, Koppeschaar H, Bernaards C, Thijssen J, Verbaten R (February 2000). "Time course of effects of testosterone administration on sexual arousal in women". Archives of General Psychiatry. 57 (2): 149–53, discussion 155–6. doi:10.1001/archpsyc.57.2.149. PMID10665617.
12Bolour S, Braunstein G (2005). "Testosterone therapy in women: a review". International Journal of Impotence Research. 17 (5): 399–408. doi:10.1038/sj.ijir.3901334. PMID15889125.
12van Anders SM, Watson NV (July 2006). "Relationship status and testosterone in North American heterosexual and non-heterosexual men and women: cross-sectional and longitudinal data". Psychoneuroendocrinology. 31 (6): 715–23. doi:10.1016/j.psyneuen.2006.01.008. hdl:2027.42/83924. PMID16621328. S2CID22477678.
123Booth A, Dabbs JM (1993). "Testosterone and Men's Marriages". Social Forces. 72 (2): 463–77. doi:10.1093/sf/72.2.463.
↑Mazur A, Michalek J (1998). "Marriage, Divorce, and Male Testosterone". Social Forces. 77 (1): 315–30. doi:10.1093/sf/77.1.315.
↑Bribiescas RG, Ellison PT, Gray PB (December 2012). "Male Life History, Reproductive Effort, and the Evolution of the Genus Homo". Current Anthropology. 53 (S6): S424 –S435. doi:10.1086/667538. S2CID83046141.
↑Armstrong TA, Boisvert DL, Wells J, Lewis RH, Cooke EM, Woeckener M, etal. (November 2022). "Testosterone, cortisol, and criminal behavior in men and women". Hormones and Behavior. 146 105260. doi:10.1016/j.yhbeh.2022.105260. PMID36122515. S2CID252285821.
↑Dabbs JM, Jurkovic GJ, Frady RL (August 1991). "Salivary testosterone and cortisol among late adolescent male offenders". Journal of Abnormal Child Psychology. 19 (4): 469–78. doi:10.1007/BF00919089. PMID1757712. S2CID647349.
↑Dabbs Jr JM, Carr TS, Frady RL, Riad JK (May 1995). "Testosterone, crime, and misbehavior among 692 male prison inmates". Personality and Individual Differences. 18 (5): 627–633. doi:10.1016/0191-8869(94)00177-T.
↑Welker KM, Lozoya E, Campbell JA, Neumann CS, Carré JM (April 2014). "Testosterone, cortisol, and psychopathic traits in men and women". Physiology & Behavior. 129: 230–6. doi:10.1016/j.physbeh.2014.02.057. PMID24631306. S2CID23683791.
↑Butovskaya M, Burkova V, Karelin D, Fink B (October 1, 2015). "Digit ratio (2D:4D), aggression, and dominance in the Hadza and the Datoga of Tanzania". American Journal of Human Biology. 27 (5): 620–627. doi:10.1002/ajhb.22718. PMID25824265. S2CID205303673.
↑Joyce CW, Kelly JC, Chan JC, Colgan G, O'Briain D, Mc Cabe JP, etal. (November 2013). "Second to fourth digit ratio confirms aggressive tendencies in patients with boxers fractures". Injury. 44 (11): 1636–1639. doi:10.1016/j.injury.2013.07.018. PMID23972912.
↑Klinesmith J, Kasser T, McAndrew FT (July 2006). "Guns, testosterone, and aggression: an experimental test of a mediational hypothesis". Psychological Science. 17 (7): 568–571. doi:10.1111/j.1467-9280.2006.01745.x. PMID16866740. S2CID33952211.
↑von der PB, Sarkola T, Seppa K, Eriksson CJ (September 2002). "Testosterone, 5 alpha-dihydrotestosterone and cortisol in men with and without alcohol-related aggression". Journal of Studies on Alcohol. 63 (5): 518–26. doi:10.15288/jsa.2002.63.518. PMID12380846.
↑Goldman D, Lappalainen J, Ozaki N. Direct analysis of candidate genes in impulsive disorders. In: Bock G, Goode J, eds. Genetics of Criminal and Antisocial Behaviour. Ciba Foundation Symposium 194. Chichester: John Wiley & Sons; 1996.
↑Soma KK, Scotti MA, Newman AE, Charlier TD, Demas GE (October 2008). "Novel mechanisms for neuroendocrine regulation of aggression". Frontiers in Neuroendocrinology. 29 (4): 476–89. doi:10.1016/j.yfrne.2007.12.003. PMID18280561. S2CID32650274.
↑Soma KK, Sullivan KA, Tramontin AD, Saldanha CJ, Schlinger BA, Wingfield JC (2000). "Acute and chronic effects of an aromatase inhibitor on territorial aggression in breeding and nonbreeding male song sparrows". Journal of Comparative Physiology A. 186 (7–8): 759–69. doi:10.1007/s003590000129. PMID11016791. S2CID23990605.
↑McGinnis MY, Lumia AR, Breuer ME, Possidente B (February 2002). "Physical provocation potentiates aggression in male rats receiving anabolic androgenic steroids". Hormones and Behavior. 41 (1): 101–10. doi:10.1006/hbeh.2001.1742. PMID11863388. S2CID29969145.
↑Cunningham SK, Loughlin T, Culliton M, McKenna TJ (September 1985). "The relationship between sex steroids and sex-hormone-binding globulin in plasma in physiological and pathological conditions". Ann Clin Biochem. 22 (5): 489–97. doi:10.1177/000456328502200504. PMID4062218.
↑McPhaul MJ, Young M (September 2001). "Complexities of androgen action". Journal of the American Academy of Dermatology. 45 (3 Suppl): S87–94. doi:10.1067/mjd.2001.117429. PMID11511858.
↑Bennett NC, Gardiner RA, Hooper JD, Johnson DW, Gobe GC (2010). "Molecular cell biology of androgen receptor signalling". Int. J. Biochem. Cell Biol. 42 (6): 813–27. doi:10.1016/j.biocel.2009.11.013. PMID19931639.
↑Breiner M, Romalo G, Schweikert HU (August 1986). "Inhibition of androgen receptor binding by natural and synthetic steroids in cultured human genital skin fibroblasts". Klinische Wochenschrift. 64 (16): 732–37. doi:10.1007/BF01734339. PMID3762019. S2CID34846760.
12Gravanis A, Calogeropoulou T, Panoutsakopoulou V, Thermos K, Neophytou C, Charalampopoulos I (2012). "Neurosteroids and microneurotrophins signal through NGF receptors to induce prosurvival signaling in neuronal cells". Sci Signal. 5 (246): pt8. doi:10.1126/scisignal.2003387. PMID23074265. S2CID26914550.
↑Albayrak Y, Hashimoto K (2017). "Sigma-1 Receptor Agonists and Their Clinical Implications in Neuropsychiatric Disorders". Sigma Receptors: Their Role in Disease and as Therapeutic Targets. Advances in Experimental Medicine and Biology. Vol.964. Springer. pp.153–161. doi:10.1007/978-3-319-50174-1_11. ISBN978-3-319-50172-7. PMID28315270.
↑Payne AH, O'Shaughnessy P (1996). "Structure, function, and regulation of steroidogenic enzymes in the Leydig cell". In Payne AH, Hardy MP, Russell LD (eds.). Leydig Cell. Vienna [Il]: Cache River Press. pp.260–85. ISBN978-0-9627422-7-9.
↑Swerdloff RS, Wang C, Bhasin S (April 1992). "Developments in the control of testicular function". Baillière's Clinical Endocrinology and Metabolism. 6 (2): 451–83. doi:10.1016/S0950-351X(05)80158-2. PMID1377467.
↑Liverman CT, Blazer DG, etal. (Institute of Medicine (US) Committee on Assessing the Need for Clinical Trials of Testosterone Replacement Therapy) (January 1, 2004). "Introduction". Testosterone and Aging: Clinical Research Directions. National Academies Press (US). doi:10.17226/10852. ISBN978-0-309-09063-6. PMID25009850. Archived from the original on January 10, 2016. Retrieved September 26, 2016– via www.ncbi.nlm.nih.gov.
↑Huhtaniemi IT (2014). "Andropause--lessons from the European Male Ageing Study". Annales d'Endocrinologie. 75 (2): 128–31. doi:10.1016/j.ando.2014.03.005. PMID24793989.
↑Vingren JL, Kraemer WJ, Ratamess NA, Anderson JM, Volek JS, Maresh CM (2010). "Testosterone physiology in resistance exercise and training: the up-stream regulatory elements". Sports Medicine. 40 (12): 1037–53. doi:10.2165/11536910-000000000-00000. PMID21058750. S2CID11683565.
↑Hulmi JJ, Ahtiainen JP, Selänne H, Volek JS, Häkkinen K, Kovanen V, etal. (May 2008). "Androgen receptors and testosterone in men—effects of protein ingestion, resistance exercise and fiber type". The Journal of Steroid Biochemistry and Molecular Biology. 110 (1–2): 130–37. doi:10.1016/j.jsbmb.2008.03.030. PMID18455389. S2CID26280370.
↑Hackney AC, Moore AW, Brownlee KK (2005). "Testosterone and endurance exercise: development of the "exercise-hypogonadal male condition"". Acta Physiologica Hungarica. 92 (2): 121–37. doi:10.1556/APhysiol.92.2005.2.3. PMID16268050.
↑Andersen ML, Tufik S (October 2008). "The effects of testosterone on sleep and sleep-disordered breathing in men: its bidirectional interaction with erectile function". Sleep Medicine Reviews. 12 (5): 365–79. doi:10.1016/j.smrv.2007.12.003. PMID18519168.
↑Akdoğan M, Tamer MN, Cüre E, Cüre MC, Köroğlu BK, Delibaş N (May 2007). "Effect of spearmint (Mentha spicata Labiatae) teas on androgen levels in women with hirsutism". Phytotherapy Research. 21 (5): 444–47. doi:10.1002/ptr.2074. PMID17310494. S2CID21961390.
↑Kumar V, Kural MR, Pereira BM, Roy P (December 2008). "Spearmint induced hypothalamic oxidative stress and testicular anti-androgenicity in male rats - altered levels of gene expression, enzymes and hormones". Food and Chemical Toxicology. 46 (12): 3563–70. doi:10.1016/j.fct.2008.08.027. PMID18804513.
↑Grant P (February 2010). "Spearmint herbal tea has significant anti-androgen effects in polycystic ovarian syndrome. A randomized controlled trial". Phytotherapy Research. 24 (2): 186–88. doi:10.1002/ptr.2900. PMID19585478. S2CID206425734.
↑Armanini D, Fiore C, Mattarello MJ, Bielenberg J, Palermo M (September 2002). "History of the endocrine effects of licorice". Experimental and Clinical Endocrinology & Diabetes. 110 (6): 257–61. doi:10.1055/s-2002-34587. PMID12373628.
↑Cumming DC, Wall SR (November 1985). "Non-sex hormone-binding globulin-bound testosterone as a marker for hyperandrogenism". The Journal of Clinical Endocrinology & Metabolism. 61 (5): 873–6. doi:10.1210/jcem-61-5-873. PMID4044776.
1234Wecker L, Watts S, Faingold C, Dunaway G, Crespo L (April 1, 2009). Brody's Human Pharmacology. Elsevier Health Sciences. pp.468–469. ISBN978-0-323-07575-6. Archived from the original on January 11, 2023. Retrieved November 3, 2016.
↑Trager L (1977). Steroidhormone: Biosynthese, Stoffwechsel, Wirkung (in German). Springer-Verlag. p.349. ISBN978-0-387-08012-3.
↑Randall VA (April 1994). "Role of 5 alpha-reductase in health and disease". Baillière's Clinical Endocrinology and Metabolism. 8 (2): 405–31. doi:10.1016/S0950-351X(05)80259-9. PMID8092979.
↑Morgentaler A (May 2017). "Andrology: Testosterone reference ranges and diagnosis of testosterone deficiency". Nature Reviews. Urology. 14 (5): 263–264. doi:10.1038/nrurol.2017.35. PMID28266512. S2CID29122481.
↑Morgentaler A, Khera M, Maggi M, Zitzmann M (July 2014). "Commentary: Who is a candidate for testosterone therapy? A synthesis of international expert opinions". The Journal of Sexual Medicine. 11 (7): 1636–1645. doi:10.1111/jsm.12546. PMID24797325.
12Steinberger E, Ayala C, Hsi B, Smith KD, Rodriguez-Rigau LJ, Weidman ER, etal. (1998). "Utilization of commercial laboratory results in management of hyperandrogenism in women". Endocrine Practice. 4 (1): 1–10. doi:10.4158/EP.4.1.1. PMID15251757.
↑"RCSB PDB - 1D2S". Crystal Structure of the N-Terminal Laminin G-Like Domain of SHBG in Complex with Dihydrotestosterone. Archived from the original on June 28, 2021. Retrieved February 19, 2019.
↑Nassar GW, Leslie S (2024). "Physiology, Testosterone". StatPearls. Treasure Island (FL): StatPearls Publishing. PMID30252384. Archived from the original on October 2, 2023. Retrieved March 3, 2024.
↑Berthold AA (1849). "Transplantation der Hoden" [Transplantation of testis]. Arch. Anat. Physiol. Wiss. (in German). 16: 42–46.
↑David KG, Dingemanse E, Freud JL (May 1935). "Über krystallinisches mannliches Hormon aus Hoden (Testosteron) wirksamer als aus harn oder aus Cholesterin bereitetes Androsteron" [On crystalline male hormone from testicles (testosterone) effective as from urine or from cholesterol]. Hoppe-Seyler's Z Physiol Chem (in German). 233 (5–6): 281–83. doi:10.1515/bchm2.1935.233.5-6.281.
↑Butenandt A, Hanisch G (1935). "Umwandlung des Dehydroandrosterons in Androstendiol und Testosterone; ein Weg zur Darstellung des Testosterons aus Cholestrin" [About Testosterone. Conversion of Dehydro-androsterons into androstendiol and testosterone; a way for the structure assignment of testosterone from cholesterol]. Hoppe-Seyler's Z Physiol Chem (in German). 237 (2): 89–97. doi:10.1515/bchm2.1935.237.1-3.89.
↑Butenandt A, Hanisch G (1935). "Uber die Umwandlung des Dehydroandrosterons in Androstenol-(17)-one-(3) (Testosterone); um Weg zur Darstellung des Testosterons auf Cholesterin (Vorlauf Mitteilung). [The conversion of dehydroandrosterone into androstenol-(17)-one-3 (testosterone); a method for the production of testosterone from cholesterol (preliminary communication)]". Chemische Berichte (in German). 68 (9): 1859–62. doi:10.1002/cber.19350680937.
↑Ruzicka L, Wettstein A (1935). "Uber die kristallinische Herstellung des Testikelhormons, Testosteron (Androsten-3-ol-17-ol) [The crystalline production of the testicle hormone, testosterone (Androsten-3-ol-17-ol)]". Helvetica Chimica Acta (in German). 18: 1264–75. doi:10.1002/hlca.193501801176.
↑Kenyon AT, Knowlton K, Sandiford I, Koch FC, Lotwin, G (February 1940). "A comparative study of the metabolic effects of testosterone propionate in normal men and women and in eunuchoidism". Endocrinology. 26 (1): 26–45. doi:10.1210/Endo-26-1-26.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.