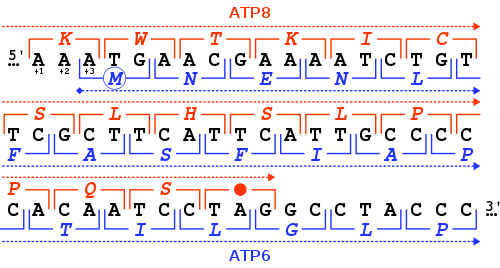
The start codon is the first codon of a messenger RNA (mRNA) transcript translated by a ribosome. The start codon always codes for methionine in eukaryotes and archaea and a N-formylmethionine (fMet) in bacteria, mitochondria and plastids.
Contents
- Decoding
- Alternative start codons
- Eukaryotes
- Bacteria
- Mitochondria
- Archaea
- Upstream start codons
- Standard genetic code
- Non-methionine start codons
- Natural
- Engineered start codons
- See also
- References
- External links
The start codon is often preceded by a 5' untranslated region (5' UTR). In prokaryotes this includes the ribosome binding site.