This article is missing information about genetic and architectural classification (ConoServer and PMC4278219).(April 2019) |
| Alpha conotoxin precursor | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 α-Conotoxin PnIB from C. pennaceus, disulfide bonds shown in yellow. From the University of Michigan's Orientations of Proteins in Membranes database, PDB: 1AKG . | |||||||||
| Identifiers | |||||||||
| Symbol | Toxin_8 | ||||||||
| Pfam | PF07365 | ||||||||
| InterPro | IPR009958 | ||||||||
| PROSITE | PDOC60004 | ||||||||
| SCOP2 | 1mii / SCOPe / SUPFAM | ||||||||
| OPM superfamily | 148 | ||||||||
| OPM protein | 1akg | ||||||||
| |||||||||
| Omega conotoxin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
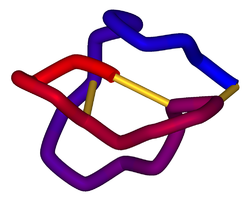 Schematic diagram of the three-dimensional structure of ω-conotoxin MVIIA (ziconotide). Disulfide bonds are shown in gold. From PDB: 1DW5 . | |||||||||
| Identifiers | |||||||||
| Symbol | Conotoxin | ||||||||
| Pfam | PF02950 | ||||||||
| InterPro | IPR004214 | ||||||||
| SCOP2 | 2cco / SCOPe / SUPFAM | ||||||||
| OPM superfamily | 112 | ||||||||
| OPM protein | 1fyg | ||||||||
| |||||||||
A conotoxin is one of a group of neurotoxic peptides isolated from the venom of the marine cone snail, genus Conus .
Contents
- Hypervariability
- Disulfide connectivities
- Types and biological activities
- Alpha
- Delta, kappa, and omega
- Mu
- See also
- References
- External links
Conotoxins, which are peptides consisting of 10 to 30 amino acid residues, typically have one or more disulfide bonds. Conotoxins have a variety of mechanisms of actions, most of which have not been determined. However, it appears that many of these peptides modulate the activity of ion channels. [1] Over the last few decades conotoxins have been the subject of pharmacological interest. [2]
