



Toll-like receptor 8 is a protein that in humans is encoded by the TLR8 gene. [5] TLR8 has also been designated as CD288 (cluster of differentiation 288). It is a member of the toll-like receptor (TLR) family.
Toll-like receptor 8 is a protein that in humans is encoded by the TLR8 gene. [5] TLR8 has also been designated as CD288 (cluster of differentiation 288). It is a member of the toll-like receptor (TLR) family.
TLR8 seems to function differently in humans and mice. Until recently, TLR8 was believed to be nonfunctional in mice, but it seems to counteract TLR7 activity [6] [7]
The TLR family plays a fundamental role in pathogen recognition and activation of innate immunity. TLRs are highly conserved from Drosophila to humans and share structural and functional similarities. They recognize pathogen-associated molecular patterns (PAMPs) that are expressed on infectious agents, and mediate the production of cytokines necessary for the development of effective immunity. The various TLRs exhibit different patterns of expression. This gene is predominantly expressed in lung and peripheral blood leukocytes, and lies in close proximity to another family member, TLR7, on chromosome X. [8] Recent research has also shown the expression of TLR8 in hippocampal interneurons, with yet unknown function. [9]
TLR8 can recognize GU-rich single-stranded RNA. [10] However, the presence of GU-rich sequences in the single-stranded RNA is not sufficient to stimulate TLR8. [11] TLR8 recognizes G-rich oligonucleotides. [12] TLR8 is activated by ssRNA and forms a dimer complex when uridine released from the degraded ssRNA binds at one active site in between the dimers and a short oligonucleotide binds to another active site on the surface of the TLR8 structure. [13]
TLR8 is an endosomal receptor that recognizes single stranded RNA (ssRNA), and can recognize ssRNA viruses such as Influenza, Sendai, and Coxsackie B viruses. TLR8 binding to the viral RNA recruits the myeloid differentiation primary response protein 88 (MyD88) and leads to activation of the transcription factor NF-κB and an antiviral response that leads to proinflammatory cytokine synthesis. [14] [15] TLR8 recognizes single-stranded RNA of viruses such as HIV and HCV. [10] [11] TLR8 is also involved in the activation of dendritic cells to produce inflammatory factors to help regulate tumor growth, so TLR8 is often used as a target in the research for therapies in treating cancers including ovarian cancer and lymphomas. [15]
Genetic variants in TLR8 has recently been linked to susceptibility to pulmonary tuberculosis. [16]

Toll-like receptors (TLRs) are a class of proteins that play a key role in the innate immune system. They are single-spanning receptors usually expressed on sentinel cells such as macrophages and dendritic cells, that recognize structurally conserved molecules derived from microbes. Once these microbes have reached physical barriers such as the skin or intestinal tract mucosa, they are recognized by TLRs, which activate immune cell responses. The TLRs include TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TLR11, TLR12, and TLR13. Humans lack genes for TLR11, TLR12 and TLR13 and mice lack a functional gene for TLR10. The receptors TLR1, TLR2, TLR4, TLR5, TLR6, and TLR10 are located on the cell membrane, whereas TLR3, TLR7, TLR8, and TLR9 are located in intracellular vesicles.
Pattern recognition receptors (PRRs) play a crucial role in the proper function of the innate immune system. PRRs are germline-encoded host sensors, which detect molecules typical for the pathogens. They are proteins expressed mainly by cells of the innate immune system, such as dendritic cells, macrophages, monocytes, neutrophils, as well as by epithelial cells, to identify two classes of molecules: pathogen-associated molecular patterns (PAMPs), which are associated with microbial pathogens, and damage-associated molecular patterns (DAMPs), which are associated with components of host's cells that are released during cell damage or death. They are also called primitive pattern recognition receptors because they evolved before other parts of the immune system, particularly before adaptive immunity. PRRs also mediate the initiation of antigen-specific adaptive immune response and release of inflammatory cytokines.
Toll-like receptor 3 (TLR3) also known as CD283 is a protein that in humans is encoded by the TLR3 gene. TLR3 is a member of the toll-like receptor family of pattern recognition receptors of the innate immune system. TLR3 recognizes double-stranded RNA in endosomes, which is a common feature of viral genomes internalised by macrophages and dendritic cells.

IRAK-4, in the IRAK family, is a protein kinase involved in signaling innate immune responses from Toll-like receptors. It also supports signaling from T-cell receptors. IRAK4 contains domain structures which are similar to those of IRAK1, IRAK2, IRAKM and Pelle. IRAK4 is unique compared to IRAK1, IRAK2 and IRAKM in that it functions upstream of the other IRAKs, but is more similar to Pelle in this trait. IRAK4 has important clinical applications.

Myeloid differentiation primary response 88 (MYD88) is a protein that, in humans, is encoded by the MYD88 gene. originally discovered in the laboratory of Dan A. Liebermann as a Myeloid differentiation primary response gene.

Toll-like receptor 2 also known as TLR2 is a protein that in humans is encoded by the TLR2 gene. TLR2 has also been designated as CD282. TLR2 is one of the toll-like receptors and plays a role in the immune system. TLR2 is a membrane protein, a receptor, which is expressed on the surface of certain cells and recognizes foreign substances and passes on appropriate signals to the cells of the immune system.

TIR domain containing adaptor molecule 1 is an adapter in responding to activation of toll-like receptors (TLRs). It mediates the rather delayed cascade of two TLR-associated signaling cascades, where the other one is dependent upon a MyD88 adapter.

Toll-like receptor 7, also known as TLR7, is a protein that in humans is encoded by the TLR7 gene. Orthologs are found in mammals and birds. It is a member of the toll-like receptor (TLR) family and detects single stranded RNA.

Toll-like receptor 5, also known as TLR5, is a protein which in humans is encoded by the TLR5 gene. It is a member of the toll-like receptor (TLR) family. TLR5 is known to recognize bacterial flagellin from invading mobile bacteria. It has been shown to be involved in the onset of many diseases, which includes Inflammatory bowel disease. Recent studies have also shown that malfunctioning of TLR5 is likely related to rheumatoid arthritis, osteoclastogenesis, and bone loss. Abnormal TLR5 functioning is related to the onset of gastric, cervical, endometrial and ovarian cancers.

Toll-like receptor 4 (TLR4), also designated as CD284, is a key activator of the innate immune response and plays a central role in the fight against bacterial infections. TLR4 is a transmembrane protein of approximately 95 kDa that is encoded by the TLR4 gene.

Toll-like receptor 6 is a protein that in humans is encoded by the TLR6 gene. TLR6 is a transmembrane protein, member of toll-like receptor family, which belongs to the pattern recognition receptor (PRR) family. TLR6 acts in a heterodimer form with toll-like receptor 2 (TLR2). Its ligands include multiple diacyl lipopeptides derived from gram-positive bacteria and mycoplasma and several fungal cell wall saccharides. After dimerizing with TLR2, the NF-κB intracellular signalling pathway is activated, leading to a pro-inflammatory cytokine production and activation of innate immune response. TLR6 has also been designated as CD286.

Toll-like receptor 9 is a protein that in humans is encoded by the TLR9 gene. TLR9 has also been designated as CD289. It is a member of the toll-like receptor (TLR) family. TLR9 is an important receptor expressed in immune system cells including dendritic cells, macrophages, natural killer cells, and other antigen presenting cells. TLR9 is expressed on endosomes internalized from the plasma membrane, binds DNA, and triggers signaling cascades that lead to a pro-inflammatory cytokine response. Cancer, infection, and tissue damage can all modulate TLR9 expression and activation. TLR9 is also an important factor in autoimmune diseases, and there is active research into synthetic TLR9 agonists and antagonists that help regulate autoimmune inflammation.

Toll-like receptor 10 is a protein that in humans is encoded by the TLR10 gene. TLR10 has also been designated as CD290 . TLR10 has not been extensively studied because it is a pseudogene in mice, though all other mammalian species contain an intact copy of the TLR10 gene. Unlike other TLRs, TLR10 does not activate the immune system and has instead been shown to suppress inflammatory signaling on primary human cells. This makes TLR10 unique among the TLR family. TLR10 was thought to be an "orphan" receptor, however, recent studies have identified ligands for TLR10 and these include HIV-gp41. Ligands for TLR2 are potential ligands for TLR10.

RIG-I is a cytosolic pattern recognition receptor (PRR) that can mediate induction of a type-I interferon (IFN1) response. RIG-I is an essential molecule in the innate immune system for recognizing cells that have been infected with a virus. These viruses can include West Nile virus, Japanese Encephalitis virus, influenza A, Sendai virus, flavivirus, and coronaviruses.

Interferon alpha-1 is a protein that in humans is encoded by the IFNA1 gene.

Mitochondrial antiviral-signaling protein (MAVS) is a protein that is essential for antiviral innate immunity. MAVS is located in the outer membrane of the mitochondria, peroxisomes, and mitochondrial-associated endoplasmic reticulum membrane (MAM). Upon viral infection, a group of cytosolic proteins will detect the presence of the virus and bind to MAVS, thereby activating MAVS. The activation of MAVS leads the virally infected cell to secrete cytokines. This induces an immune response which kills the host's virally infected cells, resulting in clearance of the virus.

MDA5 is a RIG-I-like receptor dsRNA helicase enzyme that is encoded by the IFIH1 gene in humans. MDA5 is part of the RIG-I-like receptor (RLR) family, which also includes RIG-I and LGP2, and functions as a pattern recognition receptor capable of detecting viruses. It is generally believed that MDA5 recognizes double stranded RNA (dsRNA) over 2000nts in length, however it has been shown that whilst MDA5 can detect and bind to cytoplasmic dsRNA, it is also activated by a high molecular weight RNA complex composed of ssRNA and dsRNA. For many viruses, effective MDA5-mediated antiviral responses are dependent on functionally active LGP2. The signaling cascades in MDA5 is initiated via CARD domain. Some observations made in cancer cells show that MDA5 also interacts with cellular RNA is able to induce an autoinflammatory response.
An interferon-stimulated gene (ISG) is a gene that can be expressed in response to stimulation by interferon. Interferons bind to receptors on the surface of a cell, initiating protein signaling pathways within the cell. This interaction leads to the expression of a subset of genes involved in the innate immune system response. ISGs are commonly expressed in response to viral infection, but also during bacterial infection and in the presence of parasites. It's currently estimated that 10% of the human genome is regulated by interferons (IFNs). Interferon stimulated genes can act as an initial response to pathogen invasion, slowing down viral replication and increasing expression of immune signaling complexes. There are three known types of interferon. With approximately 450 genes highly expressed in response to interferon type I. Type I interferon consists of INF-α, INF-β, INF-ω and is expressed in response to viral infection. ISGs induced by type I interferon are associated with viral replication suppression and increase expression of immune signaling proteins. Type II interferon consists only of INF-γ and is associated with controlling intracellular pathogens and tumor suppressor genes. Type III interferon consists of INF-λ and is associated with viral immune response and is key in anti-fungal neutrophil response.
The interleukin-1 receptor (IL-1R) associated kinase (IRAK) family plays a crucial role in the protective response to pathogens introduced into the human body by inducing acute inflammation followed by additional adaptive immune responses. IRAKs are essential components of the Interleukin-1 receptor signaling pathway and some Toll-like receptor signaling pathways. Toll-like receptors (TLRs) detect microorganisms by recognizing specific pathogen-associated molecular patterns (PAMPs) and IL-1R family members respond the interleukin-1 (IL-1) family cytokines. These receptors initiate an intracellular signaling cascade through adaptor proteins, primarily, MyD88. This is followed by the activation of IRAKs. TLRs and IL-1R members have a highly conserved amino acid sequence in their cytoplasmic domain called the Toll/Interleukin-1 (TIR) domain. The elicitation of different TLRs/IL-1Rs results in similar signaling cascades due to their homologous TIR motif leading to the activation of mitogen-activated protein kinases (MAPKs) and the IκB kinase (IKK) complex, which initiates a nuclear factor-κB (NF-κB) and AP-1-dependent transcriptional response of pro-inflammatory genes. Understanding the key players and their roles in the TLR/IL-1R pathway is important because the presence of mutations causing the abnormal regulation of Toll/IL-1R signaling leading to a variety of acute inflammatory and autoimmune diseases.
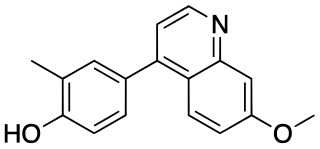
CU-CPT9a is a drug which acts as a potent and selective antagonist of Toll-like receptor 8 (TLR8), with an IC50 of 0.5nM. Activation of toll-like receptors triggers release of cytokines and other signalling factors, leading to inflammation. This is an essential part of the immune system's response to infection, but chronic activation of TLR signalling is thought to be involved in various inflammatory and autoimmune disorders. CU-CPT9a has immunosuppressant properties, and may have applications in the treatment of autoimmune disorders, as well as being used in scientific research into the function of TLR8.