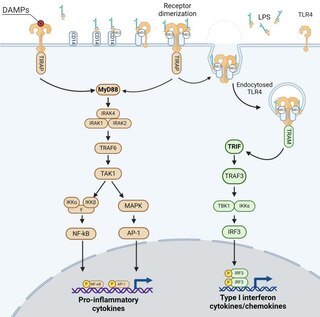
TLR4 has been reported to play both friend and foe in a variety of human diseases, such as bacterial infections and cancers. This dual role of TLR4 depends on the intensity, duration and site (surface or endosome) of its activation, its polymorphism and the balance of activation of signaling pathways (MyD88 vs. TRIF).
Cancer
The role of the TLR4 in the control of cancer progression and in cancer therapy is well documented.
Stimulation of TLR4 by natural derivatives and LPS is well known to induce potent antitumor activity. This anti-tumor activity is linked to the ability of LPS to stimulate innate immunity via TLR4, resulting in the production of pro-inflammatory cytokines and type 1 interferons, and the indirect generation of adaptive anti-tumor responses. [64] [65]
The first clues about the efficacy of TLR4 agonists like LPS in cancer immunotherapy was found in the 19th centuries, when bacterial infections were found to induce tumor regressions. [66] Later, Dr William Coley showed the therapeutic efficacy of a mixed bacterial vaccine, so-called "Coley's toxin", to human cancer. [67] Since then, a number of developments have been made in the treatment or prevention of cancer using bacterial mixtures strongly activating TLR4 due to LPS content. The antituberculosis vaccine Bacillus Calmette–Guérin (BCG) was approved by the Federal Drug Administration (FDA) in 1990 for the local treatment of superficial bladder cancer. BCG promotes dendritic cell maturation, and this effect is TLR4 (as well as TLR2) dependent. [68] There are also reports on the treatment of oral squamous cell carcinoma, gastric , Head-and-neck and cervical cancers with lyophilized streptococcal preparation OK-432 (Picibanil). [69] The mechanism of action of OK-432 involves TLR4 activation, since OKA-432 does not inhibit tumor growth on TLR4 knockouts as it does on wild-type mice. [70]
Purified LPS also showed potent anti-tumor efficacy as systemic therapeutic agents in several tumor models. [71] [72] In the 90's, clinical trials evaluating the intravenous administration of LPS to patients with cancer provided positive results including several cases of disease stabilization and partial responses. However, limiting toxicities at doses in the ng/kg range has been reported which are too low to obtain significant antitumor effects. [73]
Subsequently, detoxified TLR4 agonists (LPS derivatives) have been produced and evaluated in the clinic. This includes the MPL, a chemically modified LPS which was the first TLR4 agonist to be approved and commercialized by GSK in 5 human vaccines (HPV, Zoster, Hepatitis B, Malaria, RSV). MPL was investigated as an adjuvant for curative anti-tumor vaccines, with the approval of Melacine in Canada for the treatment of patients with malignant melanoma. [74] Synthetic LPS derivatives based on dephosphorylated lipid A moiety structures were also developed and confirmed potent adjuvant and antitumor activities as therapeutic agents. In particular, the intratumoral administration of Glucopyranosyl Lipid Adjuvant (GLA-SE/G100), a synthetic detoxified analog of lipid A formulated in a stable emulsion, showed anti-tumor immune responses and tumor regression in patients with Merkel cell carcinoma, [75] and potent adjuvant activity in phase 2 trials in combination with pembrolizumab in patients with follicular lymphoma. [76] [77]
Besides the recognized anti-tumor efficacy of TLR4 activation by LPS, some studies suggest that TLR4 may also contribute to the development of some cancers, (prostate, liver, breast and lung cancers) and may contribute to resistance to paclitaxel chemotherapy in breast cancer. [78] Some clinical studies also suggested a potential correlation between TLR4 expression on tumor cells and tumor progression. However, no such effect was reported in the numerous clinical studies conducted with natural LPS or LPS derivatives. On the contrary, in phase 2 studies with GLA, a positive association between baseline TLR4 expression in tumors and the increase of overall response rates has been reported. [77]
The potential impact of TLR4 on the progression of some cancers was associated with the excessive production of pro-inflammatory cytokines via activation of the TLR4-MyD88/NF-kB signaling pathway. [79] [80] [81] Several studies showed that this is mediated by the misuse of DAMP signaling by tumor cells. [12] [82] [14]
Many DAMPs are released by dying or necrotic tumor cells and present during cancer progression. DAMPs released from tumor cells can directly activate tumor-expressed TLR4 that induce chemoresistance, migration, invasion, and metastasis. Furthermore, DAMP-induced chronic inflammation in the tumor microenvironment causes an increase in immunosuppressive populations, such as M2 macrophages, myeloid-derived suppressor cells (MDSCs), and regulatory T cells (Tregs). [12] DAMPs, such as HMGB1, S100 proteins, and heat shock proteins (HSPs), were found to strongly activate inflammatory pathways and release IL-1, IL-6, LT-β, IFN-γ, TNF, and transforming growth factor (TGF)-β promoting inflammation, immunosuppression, angiogenesis, and tumor cell proliferation. [11]
Several studies have evaluated the potential association of this TLR4 polymorphism with cancer risk, but the data are highly conflicting. However, some meta-analyses suggest an association of SNP D299G with gastric, viral-induced and female-specific cancers (cervix, ovary). [83]
Neurogenerative diseases
Growing evidence suggests an implication of TLR4 in the development and progression of neurogenerative disorders such as Alzheimer's disease, Parkinson's disease, and Huntington's disease. In the brain, TLR4 is expressed by neurons as well as the non-neuronal glial cells, which include microglia, astrocytes, and oligodendrocytes. TLR4 is expressed primarily by microglia, and to a lesser extent by astrocytes, oligodendrocytes, and neurons. [5] Microglia are representatives of the mononuclear phagocyte system in the brain, and TLR4 activation regulates some of their functions, such as phagocytic activity. [84] [13]
Activation of microglial TLR4 has been suggested to protect against or slow the development of neurodegenerative diseases, notably by enhancing the clearance of neurotoxic proteins such as Aβ and its aggregates, thanks to increased phagocytic and autophagic activity. [85]
However, chronic TLR4 activation is believed to be associated with glia-mediated neuronal death due to excessive secretion of pro-inflammatory cytotoxins leading to neuroinflammation, a key factor in the development of many neurodegenerative diseases. [86] [87] In the brain, TLR4 can be activated by various endogenous DAMPs in addition to pathology-associated proteins such as aggregates of amyloid-βpeptides (Aβ) or α-synuclein. [88] All these structures bind TLR4 and activate downstream signaling pathways in glia, inducing secretion of reactive oxygen species (ROS) and proinflammatory cytokines such as IL-1β and TNF-α, which can lead to damage and death of neurons. [86] [89] [90] Neuronal death is accompanied by the release of DAMPs into the extracellular space, which can then further activate TLR4, aggravating neuroinflammation. [91] In patients with Alzheimer's disease (AD), the levels of circulating DAMPs like HMGB1 and soluble RAGE, are significantly elevated, which was correlated with the levels of amyloid beta. [92] In AD patients, the serum levels of S100B are also intimately related to the severity of the disease. [93] The role of the HMGB1-TLR4 axis is very important in the pathogenesis of Parkinson's disease (PD). The serum HMGB1 and TLR4 protein levels were significantly elevated in PD patients and correlated with the PD stages. [94]
Targeting TLR4 with agonists or antagonists, or modulating its downstream signaling pathways, may have a therapeutic potential in treating neurodegenerative diseases. [95] TLR4-specific antagonists could suppress neuroinflammation by reducing overproduction of inflammatory mediators and cytotoxins by glia. However, TLR4 antagonists could have adverse CNS effects by inhibiting phagocytosis by glia, reducing protein clearance, and interfering with myelination. [96] Some studies showed that selective TLR4 agonists could be beneficial by upregulating the phagocytic activity of microglia, leading to enhanced clearance of damaged tissue and abnormal protein aggregates associated with several different CNS diseases. Repeated injections of MPL, at doses that are nonpyrogenic, were found to significantly improved AD-related pathology mice. [97] MPL led to a significant reduction in Aβ load in the brain, as well as enhanced cognitive function. MPL induced a potent phagocytic response by microglia while triggering a moderate inflammatory reaction. However, adverse effects can be caused by TLR 4 agonists inducing secretion of inflammatory mediators. Studies therefore suggested that TLR4 agonists that selectively activate the TRIF signaling pathway could be highly beneficial in the treatment of neurodegenerative disorders by increasing glial cell phagocytic activity without significantly increasing glial cytokines and cytotoxins. [96]