CD14 exists in two forms, one anchored to the membrane by a glycosylphosphatidylinositol (GPI) tail (mCD14), the other a soluble form (sCD14). Soluble CD14 either appears after shedding of mCD14 (48 kDa) or is directly secreted from intracellular vesicles (56 kDa).[7]
The x-ray crystal structure of human CD14 reveals a monomeric, bent solenoid structure containing a hydrophobic amino-terminal pocket.[8]
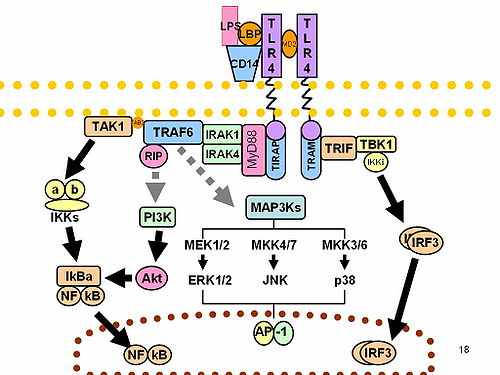
CD14 acts as a co-receptor (along with the Toll-like receptorTLR 4 and MD-2) for the detection of bacterial lipopolysaccharide (LPS).[9][10] CD14 can bind LPS only in the presence of lipopolysaccharide-binding protein (LBP). Although LPS is considered its main ligand, CD14 also recognizes other pathogen-associated molecular patterns such as lipoteichoic acid.[11] Cluster of differentiation CD14 is a receptor for a very wide range of microbial products including lipopolysaccharide (released from Gram-negative bacteria), peptidoglycans, and lipoteichoic acid (constituents of Gram-positive bacteria).[12]
CD14 is expressed mainly by macrophages and (at 10-times lesser extent) by neutrophils. It is also expressed by dendritic cells. The soluble form of the receptor (sCD14) is secreted by the liver and monocytes and is sufficient in low concentrations to confer LPS-responsiveness to cells not expressing CD14. mCD14 and sCD14 are also present on enterocytes.[13] sCD14 is also present in human milk, where it is believed to regulate microbial growth in the infant gut.
↑"Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
↑"Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
↑Setoguchi M, Nasu N, Yoshida S, Higuchi Y, Akizuki S, Yamamoto S (July 1989). "Mouse and human CD14 (myeloid cell-specific leucine-rich glycoprotein) primary structure deduced from cDNA clones". Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression. 1008 (2): 213–222. doi:10.1016/0167-4781(80)90012-3. PMID2472171.
↑Kirkland TN, Viriyakosol S (1998). "Structure-function analysis of soluble and membrane-bound CD14". Progress in Clinical and Biological Research. 397: 79–87. PMID9575549.
↑Kitchens RL (2000). "Role of CD14 in cellular recognition of bacterial lipopolysaccharides". Chemical Immunology. Chemical Immunology and Allergy. 74: 61–82. doi:10.1159/000058750. ISBN3-8055-6917-3. PMID10608082.
↑Tapping RI, Tobias PS (2000). "Soluble CD14-mediated cellular responses to lipopolysaccharide". Chemical Immunology. Chemical Immunology and Allergy. 74: 108–121. doi:10.1159/000058751. ISBN3-8055-6917-3. PMID10608084.
↑Yu B, Wright SD (1995). "LPS-dependent interaction of Mac-2-binding protein with immobilized CD14". Journal of Inflammation. 45 (2): 115–125. PMID7583357.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.