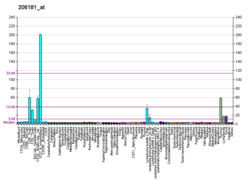
The measles virus (MV), with scientific name Morbillivirus hominis, is a single-stranded, negative-sense, enveloped, non-segmented RNA virus of the genus Morbillivirus within the family Paramyxoviridae. It is the cause of measles. Humans are the natural hosts of the virus; no animal reservoirs are known to exist.

DC-SIGN also known as CD209 is a protein which in humans is encoded by the CD209 gene.

Cluster of differentiation 40, CD40 is a type I transmembrane protein found on antigen-presenting cells and is required for their activation. The binding of CD154 (CD40L) on TH cells to CD40 activates antigen presenting cells and induces a variety of downstream effects.

Complement receptor type 2 (CR2), also known as complement C3d receptor, Epstein–Barr virus receptor, and CD21, is a protein that in humans is encoded by the CR2 gene.

Antigen presentation is a vital immune process that is essential for T cell immune response triggering. Because T cells recognize only fragmented antigens displayed on cell surfaces, antigen processing must occur before the antigen fragment can be recognized by a T-cell receptor. Specifically, the fragment, bound to the major histocompatibility complex (MHC), is transported to the surface of the antigen-presenting cell, a process known as presentation. If there has been an infection with viruses or bacteria, the antigen-presenting cell will present an endogenous or exogenous peptide fragment derived from the antigen by MHC molecules. There are two types of MHC molecules which differ in the behaviour of the antigens: MHC class I molecules (MHC-I) bind peptides from the cell cytosol, while peptides generated in the endocytic vesicles after internalisation are bound to MHC class II (MHC-II). Cellular membranes separate these two cellular environments - intracellular and extracellular. Each T cell can only recognize tens to hundreds of copies of a unique sequence of a single peptide among thousands of other peptides presented on the same cell, because an MHC molecule in one cell can bind to quite a large range of peptides. Predicting which antigens will be presented to the immune system by a certain MHC/HLA type is difficult, but the technology involved is improving.

Cluster of Differentiation 86 is a protein constitutively expressed on dendritic cells, Langerhans cells, macrophages, B-cells, and on other antigen-presenting cells. Along with CD80, CD86 provides costimulatory signals necessary for T cell activation and survival. Depending on the ligand bound, CD86 can signal for self-regulation and cell-cell association, or for attenuation of regulation and cell-cell disassociation.
NKG2 also known as CD159 is a receptor for natural killer cells. There are 7 NKG2 types: A, B, C, D, E, F and H. NKG2D is an activating receptor on the NK cell surface. NKG2A dimerizes with CD94 to make an inhibitory receptor (CD94/NKG2).

C-C chemokine receptor type 7 is a protein that in humans is encoded by the CCR7 gene. Two ligands have been identified for this receptor: the chemokines ligand 19 (CCL19/ELC) and ligand 21 (CCL21). The ligands have similar affinity for the receptor, though CCL19 has been shown to induce internalisation of CCR7 and desensitisation of the cell to CCL19/CCL21 signals. CCR7 is a transmembrane protein with 7 transmembrane domains, which is coupled with heterotrimeric G proteins, which transduce the signal downstream through various signalling cascades. The main function of the receptor is to guide immune cells to immune organs by detecting specific chemokines, which these tissues secrete.

SH2 domain–containing protein 1A is a protein that in humans is encoded by the SH2D1A gene. It is often called SLAM-associated protein, where "SLAM" refers to signaling lymphocytic activation molecules. It is a SH2 domain–containing molecule that plays a role in SLAM signaling. A putative function is as an adaptor for Fyn and competitor of phosphatases, leading to modulation of SLAM family function. SAP has been implicated in autoimmunity, and a mutation of it is associated with X-linked lymphoproliferative disease. At least 32 disease-causing mutations in this gene have been discovered.

CD244 also known as 2B4 or SLAMF4 is a protein that in humans is encoded by the CD244 gene.

Natural cytotoxicity triggering receptor 3 is a protein that in humans is encoded by the NCR3 gene. NCR3 has also been designated as CD337 and as NKp30. NCR3 belongs to the family of NCR membrane receptors together with NCR1 (NKp46) and NCR2 (NKp44).

CD84 is a human protein encoded by the CD84 gene.

CD226, PTA1 or DNAM-1 is a ~65 kDa immunoglobulin-like transmembrane glycoprotein expressed on the surface of natural killer cells, NK T cell, B cells, dendritic cells, hematopoietic precursor cells, platelets, monocytes and T cells.

SLAM family member 6 is a protein that in humans is encoded by the SLAMF6 gene.

B- and T-lymphocyte attenuator or BTLA is a protein that belongs to the CD28 immunoglobulin superfamily (IgSF) which is encoded by the BTLA gene located on the 3rd human chromosome. BTLA was first discovered in 2003 as an inhibitor of Th1 expansion and it became the 3rd member of the CD28 IgSF. However, its discovered ligand herpes virus entry mediator or HVEM belongs to the tumor necrosis factor receptor superfamily (TNFRSF). This finding was surprising because until the discovery of HVEM it was believed that receptors and ligands always belong to the same family.
T-lymphocyte surface antigen Ly-9 is a protein that in humans is encoded by the LY9 gene. LY9 has also recently been designated CD229.

OX-2 membrane glycoprotein, also named CD200 is a human protein encoded by the CD200 gene. In humans, the CD200 gene is located on chromosome 3 in proximity to genes encoding the other B7 proteins CD80/CD86. In mice, the CD200 gene is located on chromosome 16.
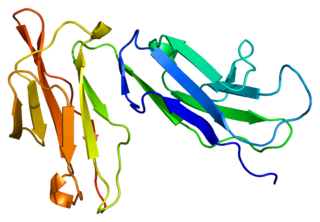
Fc fragment of IgG receptor IIb is a low affinity inhibitory receptor for the Fc region of immunoglobulin gamma (IgG). FCGR2B participates in the phagocytosis of immune complexes and in the regulation of antibody production by B lymphocytes.
Signaling lymphocytic activation molecule (SLAM) is a family of genes. Homophilic binding between SLAMs is involved in cell-to-cell adhesion during antigen presentation.
X-linked lymphoproliferative disease is a lymphoproliferative disorder, usually caused by SH2DIA gene mutations in males. XLP-positive individuals experience immune system deficiencies that render them unable to effectively respond to the Epstein-Barr virus (EBV), a common virus in humans that typically induces mild symptoms or infectious mononucleosis (IM) in patients. There are two currently known variations of the disorder, known as XLP1 and XLP2. XLP1 is estimated to occur in approximately one in every million males, while XLP2 is rarer, estimated to occur in one of every five million males. Due to therapies such as chemotherapy and stem cell transplants, the survival rate of XLP1 has increased dramatically since its discovery in the 1970s.