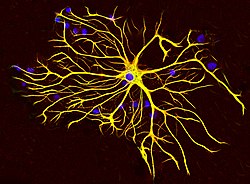
GFAP is closely related to the other three non-epithelial type III IF family members, vimentin, desmin and peripherin, which are all involved in the structure and function of the cell's cytoskeleton. GFAP is thought to help to maintain astrocytemechanical strength[14] as well as the shape of cells, but its exact function remains poorly understood, despite the number of studies using it as a cell marker. The protein was named and first isolated and characterized by Lawrence F. Eng in 1969.[15] In humans, it is located on the long arm of chromosome 17.[16]
Structure
Type III intermediate filaments contain three domains, named the head, rod and tail domains. The specific DNA sequence for the rod domain may differ between different type III intermediate filaments, but the structure of the protein is highly conserved. This rod domain coils around that of another filament to form a dimer, with the N-terminal and C-terminal of each filament aligned. Type III filaments such as GFAP are capable of forming both homodimers and heterodimers; GFAP can polymerize with other type III proteins.[17] GFAP and other type III IF proteins cannot assemble with keratins, the type I and II intermediate filaments: in cells that express both proteins, two separate intermediate filament networks form,[18] which can allow for specialization and increased variability.
To form networks, the initial GFAP dimers combine to make staggered tetramers,[19] which are the basic subunits of an intermediate filament. Since rod domains alone in vitro do not form filaments, the non-helical head and tail domains are necessary for filament formation.[17] The head and tail regions have greater variability of sequence and structure. In spite of this increased variability, the head of GFAP contains two conserved arginines and an aromatic residue that have been shown to be required for proper assembly.[20]
GFAP has been shown to play a role in mitosis by adjusting the filament network present in the cell. During mitosis, there is an increase in the amount of phosphorylated GFAP, and a movement of this modified protein to the cleavage furrow.[23] There are different sets of kinases at work; cdc2kinase acts only at the G2 phase transition, while other GFAP kinases are active at the cleavage furrow alone. This specificity of location allows for precise regulation of GFAP distribution to the daughter cells. Studies have also shown that GFAP knockout mice undergo multiple degenerative processes including abnormal myelination, white matter structure deterioration, and functional/structural impairment of the blood–brain barrier.[24] These data suggest that GFAP is necessary for many critical roles in the CNS.
GFAP is proposed to play a role in astrocyte-neuron interactions as well as cell-cell communication. In vitro, using antisense RNA, astrocytes lacking GFAP do not form the extensions usually present with neurons.[25] Studies have also shown that Purkinje cells in GFAP knockout mice do not exhibit normal structure, and these mice demonstrate deficits in conditioning experiments such as the eye-blink task.[26] Biochemical studies of GFAP have shown MgCl2 and/or calcium/calmodulin dependent phosphorylation at various serine or threonine residues by PKC and PKA[27] which are two kinases that are important for the cytoplasmic transduction of signals. These data highlight the importance of GFAP for cell-cell communication.
GFAP has also been shown to be important in repair after CNS injury. More specifically for its role in the formation of glial scars in a multitude of locations throughout the CNS including the eye[28] and brain.[29]
There are multiple disorders associated with improper GFAP regulation, and injury can cause glial cells to react in detrimental ways. Glial scarring is a consequence of several neurodegenerative conditions, as well as injury that severs neural material. The scar is formed by astrocytes interacting with fibrous tissue to re-establish the glial margins around the central injury core[32] and is partially caused by up-regulation of GFAP.[33]
Another condition directly related to GFAP is Alexander disease, a rare genetic disorder. Its symptoms include mental and physical retardation, dementia, enlargement of the brain and head, spasticity (stiffness of arms and/or legs), and seizures.[34] The cellular mechanism of the disease is the presence of cytoplasmic accumulations containing GFAP and heat shock proteins, known as Rosenthal fibers.[35] Mutations in the coding region of GFAP have been shown to contribute to the accumulation of Rosenthal fibers.[36] Some of these mutations have been proposed to be detrimental to cytoskeleton formation as well as an increase in caspase 3 activity,[37] which would lead to increased apoptosis of cells with these mutations. GFAP therefore plays an important role in the pathogenesis of Alexander disease.
The generally high abundance of GFAP in the CNS has led to a great interest in GFAP as a blood biomarker of acute injury to the brain and spinal cord in different types of disease mechanisms, such as traumatic brain injury and cerebrovascular disease.[43] After an ischemic stroke, blood levels of GFAP peak after three days and correlates strongly with infarct volume.[44] Elevated blood levels of GFAP are also found in neuroinflammatory diseases, such as multiple sclerosis and neuromyelitis optica, a disease targeting astrocytes.[43] In a study of 22 child patients undergoing extracorporeal membrane oxygenation (ECMO), children with abnormally high levels of GFAP were 13 times more likely to die and 11 times more likely to suffer brain injury than children with normal GFAP levels.[45]
Although GFAP alpha is the only isoform which is able to assemble homomerically, GFAP has 8 different isoforms which label distinct subpopulations of astrocytes in the human and rodent brain. These isoforms include GFAP kappa, GFAP +1 and the currently best researched GFAP delta. GFAP delta appears to be linked with neural stem cells (NSCs) and may be involved in migration. GFAP+1 is an antibody which labels two isoforms. Although GFAP+1 positive astrocytes are supposedly not reactive astrocytes, they have a wide variety of morphologies including processes of up to 0.95mm (seen in the human brain). The expression of GFAP+1 positive astrocytes is linked with old age and the onset of ADpathology.[48]
↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
↑ Isaacs A, Baker M, Wavrant-De Vrièze F, Hutton M (July 1998). "Determination of the gene structure of human GFAP and absence of coding region mutations associated with frontotemporal dementia with parkinsonism linked to chromosome 17". Genomics. 51 (1): 152–154. doi:10.1006/geno.1998.5360. PMID9693047.
1 2 Jacque CM, Vinner C, Kujas M, Raoul M, Racadot J, Baumann NA (January 1978). "Determination of glial fibrillary acidic protein (GFAP) in human brain tumors". Journal of the Neurological Sciences. 35 (1): 147–155. doi:10.1016/0022-510x(78)90107-7. PMID624958. S2CID10224197.
↑ Roessmann U, Velasco ME, Sindely SD, Gambetti P (October 1980). "Glial fibrillary acidic protein (GFAP) in ependymal cells during development. An immunocytochemical study". Brain Research. 200 (1): 13–21. doi:10.1016/0006-8993(80)91090-2. PMID6998542. S2CID38131934.
↑ Buniatian G, Traub P, Albinus M, Beckers G, Buchmann A, Gebhardt R, Osswald H (January 1998). "The immunoreactivity of glial fibrillary acidic protein in mesangial cells and podocytes of the glomeruli of rat kidney in vivo and in culture". Biology of the Cell. 90 (1): 53–61. doi:10.1016/s0248-4900(98)80232-3. PMID9691426. S2CID31851422.
↑ Maunoury R, Portier MM, Léonard N, McCormick D (December 1991). "Glial fibrillary acidic protein immunoreactivity in adrenocortical and Leydig cells of the Syrian golden hamster (Mesocricetus auratus)". Journal of Neuroimmunology. 35 (1–3): 119–129. doi:10.1016/0165-5728(91)90167-6. PMID1720132. S2CID3766335.
↑ Davidoff MS, Middendorff R, Köfüncü E, Müller D, Jezek D, Holstein AF (2002). "Leydig cells of the human testis possess astrocyte and oligodendrocyte marker molecules". Acta Histochemica. 104 (1): 39–49. doi:10.1078/0065-1281-00630. PMID11993850.
↑ von Koskull H (1984). "Rapid identification of glial cells in human amniotic fluid with indirect immunofluorescence". Acta Cytologica. 28 (4): 393–400. PMID6205529.
↑ Kasantikul V, Shuangshoti S (May 1989). "Positivity to glial fibrillary acidic protein in bone, cartilage, and chordoma". Journal of Surgical Oncology. 41 (1): 22–26. doi:10.1002/jso.2930410109. PMID2654484. S2CID34069861.
↑ Eng LF, Ghirnikar RS, Lee YL (October 2000). "Glial fibrillary acidic protein: GFAP-thirty-one years (1969-2000)". Neurochemical Research. 25 (9–10): 1439–1451. doi:10.1023/A:1007677003387. PMID11059815. S2CID9221868.
↑ Bongcam-Rudloff E, Nistér M, Betsholtz C, Wang JL, Stenman G, Huebner K, etal. (March 1991). "Human glial fibrillary acidic protein: complementary DNA cloning, chromosome localization, and messenger RNA expression in human glioma cell lines of various phenotypes". Cancer Research. 51 (5): 1553–1560. PMID1847665.
↑ Tardy M, Fages C, Le Prince G, Rolland B, Nunez J (1990). "Regulation of the Glial Fibrillary Acidic Protein (GFAP) and of its Encoding mRNA in the Developing Brain and in Cultured Astrocytes". Molecular Aspects of Development and Aging of the Nervous System. Advances in Experimental Medicine and Biology. Vol.265. pp.41–52. doi:10.1007/978-1-4757-5876-4_4. ISBN978-1-4757-5878-8. PMID2165732.
↑ Harrison BC, Mobley PL (January 1992). "Phosphorylation of glial fibrillary acidic protein and vimentin by cytoskeletal-associated intermediate filament protein kinase activity in astrocytes". Journal of Neurochemistry. 58 (1): 320–327. doi:10.1111/j.1471-4159.1992.tb09313.x. PMID1727439. S2CID28248825.
↑ Tuccari G, Trombetta C, Giardinelli MM, Arena F, Barresi G (1986). "Distribution of glial fibrillary acidic protein in normal and gliotic human retina". Basic and Applied Histochemistry. 30 (4): 425–432. PMID3548695.
↑ Paetau A, Elovaara I, Paasivuo R, Virtanen I, Palo J, Haltia M (1985). "Glial filaments are a major brain fraction in infantile neuronal ceroid-lipofuscinosis". Acta Neuropathologica. 65 (3–4): 190–194. doi:10.1007/bf00686997. PMID4038838. S2CID1411700.
↑ Allen A, Gulhar S, Haidari R, Martinez JP, Bekenstein J, DeLorenzo R, etal. (January 2020). "Autoimmune glial fibrillary acidic protein astrocytopathy resulting in treatment-refractory flaccid paralysis". Multiple Sclerosis and Related Disorders. 39 101924. doi:10.1016/j.msard.2019.101924. PMID31927153. S2CID210166834.
↑ Tomczak A, Su E, Tugizova M, Carlson AM, Kipp LB, Feng H, Han MH (December 2019). "A case of GFAP-astroglial autoimmunity presenting with reversible parkinsonism". Multiple Sclerosis and Related Disorders. 39 101900. doi:10.1016/j.msard.2019.101900. PMID31881522. S2CID209498996.
↑ Smith ME, Eng LF (1987). "Glial fibrillary acidic protein in chronic relapsing experimental allergic encephalomyelitis in SJL/J mice". Journal of Neuroscience Research. 18 (1): 203–208. doi:10.1002/jnr.490180129. PMID3682026. S2CID25610288.
↑ HealthLink (2007-11-25). "Alexander Disease". Medical College of Wisconsin. Archived from the original on 2007-02-24. Retrieved 2007-03-05.
↑ Brenner M, Johnson AB, Boespflug-Tanguy O, Rodriguez D, Goldman JE, Messing A (January 2001). "Mutations in GFAP, encoding glial fibrillary acidic protein, are associated with Alexander disease". Nature Genetics. 27 (1): 117–120. doi:10.1038/83679. PMID11138011. S2CID10159452.
↑ Cullen KM, Halliday GM (1994). "Chronic alcoholics have substantial glial pathology in the forebrain and diencephalon". Alcohol and Alcoholism. 2: 253–257. PMID8974344.
↑ Lopez-Egido J, Cunningham J, Berg M, Oberg K, Bongcam-Rudloff E, Gobl A (August 2002). "Menin's interaction with glial fibrillary acidic protein and vimentin suggests a role for the intermediate filament network in regulating menin activity". Experimental Cell Research. 278 (2): 175–183. doi:10.1006/excr.2002.5575. PMID12169273.
Cáceres-Marzal C, Vaquerizo J, Galán E, Fernández S (October 2006). "Early mitochondrial dysfunction in an infant with Alexander disease". Pediatric Neurology. 35 (4): 293–296. doi:10.1016/j.pediatrneurol.2006.03.010. PMID16996408.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.