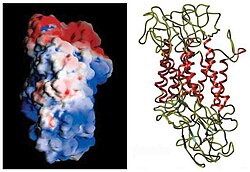
The dopamine transporter (DAT, also sodium-dependent dopamine transporter) is a membrane-spanning protein coded for in humans by the SLC6A3gene (also known as DAT1), that pumps the neurotransmitterdopamine out of the synaptic cleft back into cytosol. In the cytosol, other transporters sequester the dopamine into vesicles for storage and later release. Dopamine reuptake via DAT provides the primary mechanism through which dopamine is cleared from synapses, although there may be an exception in the prefrontal cortex, where evidence points to a possibly larger role of the norepinephrine transporter.[5]
DAT is an integral membrane protein that removes dopamine from the synaptic cleft and deposits it into surrounding cells, thus terminating the signal of the neurotransmitter. Dopamine underlies several aspects of cognition, including reward, and DAT facilitates regulation of that signal.[7]
Mechanism
DAT is a symporter that moves dopamine across the cell membrane by coupling the movement to the energetically-favorable movement of sodium ions moving from high to low concentration into the cell. DAT function requires the sequential binding and co-transport of two Na+ions and one Cl− ion with the dopamine substrate. The driving force for DAT-mediated dopamine reuptake is the ion concentration gradient generated by the plasma membrane Na+/K+ ATPase.[8]
In the most widely accepted model for monoamine transporter function, sodium ions must bind to the extracellular domain of the transporter before dopamine can bind. Once dopamine binds, the protein undergoes a conformational change, which allows both sodium and dopamine to unbind on the intracellular side of the membrane.[9]
Studies using electrophysiology and radioactive-labeled dopamine have confirmed that the dopamine transporter is similar to other monoamine transporters in that one molecule of neurotransmitter can be transported across the membrane with one or two sodium ions. Chloride ions are also needed to prevent a buildup of positive charge. These studies have also shown that transport rate and direction is totally dependent on the sodium gradient.[10]
Because of the tight coupling of the membrane potential and the sodium gradient, activity-induced changes in membrane polarity can dramatically influence transport rates. In addition, the transporter may contribute to dopamine release when the neuron depolarizes.[10]
DAT–Cav coupling
Preliminary evidence suggests that the dopamine transporter couples to L-type voltage-gated calcium channels (particularly Cav1.2 and Cav1.3), which are expressed in virtually all dopamine neurons.[11] As a result of DAT–Cav coupling, DAT substrates that produce depolarizing currents through the transporter are able to open calcium channels that are coupled to the transporter, resulting in a calcium influx in dopamine neurons.[11] This calcium influx is believed to induce CAMKII-mediated phosphorylation of the dopamine transporter as a downstream effect;[11] since DAT phosphorylation by CAMKII results in dopamine efflux in vivo, activation of transporter-coupled calcium channels is a potential mechanism by which certain drugs (e.g., amphetamine) trigger neurotransmitter release.[11]
Protein structure
The initial determination of the membrane topology of DAT was based upon hydrophobic sequence analysis and sequence similarities with the GABA transporter. These methods predicted twelve transmembrane domains (TMD) with a large extracellular loop between the third and fourth TMDs.[12] Further characterization of this protein used proteases, which digest proteins into smaller fragments, and glycosylation, which occurs only on extracellular loops, and largely verified the initial predictions of membrane topology.[13] The exact structure of the Drosophila melanogaster dopamine transporter (dDAT) was elucidated in 2013 by X-ray crystallography.[14]
Location and distribution
Pharmacodynamics of amphetamine in a dopamine neuron
Amphetamine enters the presynaptic neuron across the neuronal membrane or through DAT.[15] Once inside, it binds to TAAR1 or enters synaptic vesicles through VMAT2.[15][16][17] When amphetamine enters synaptic vesicles through VMAT2, it collapses the vesicular pH gradient, which in turn causes dopamine to be released into the cytosol (light tan-colored area) through VMAT2.[16][18] When amphetamine binds to TAAR1, it reduces the firing rate of the dopamine neuron via G protein-coupled inwardly rectifying potassium channels (GIRKs) and activates protein kinase A (PKA) and protein kinase C (PKC), which subsequently phosphorylate DAT.[15][19][20]PKA phosphorylation causes DAT to withdraw into the presynaptic neuron (internalize) and cease transport.[15]PKC-phosphorylated DAT may either operate in reverse or, like PKA-phosphorylated DAT, internalize and cease transport.[15][17] Amphetamine is also known to increase intracellular calcium, an effect which is associated with DAT phosphorylation through a CAMKIIα-dependent pathway, in turn producing dopamine efflux.[21][22]
Regional distribution of DAT has been found in areas of the brain with established dopaminergic circuitry, including the nigrostriatal, mesolimbic, and mesocortical pathways.[23] The nuclei that make up these pathways have distinct patterns of expression. Gene expression patterns in the adult mouse show high expression in the substantia nigra pars compacta.[24]
DAT in the mesocortical pathway, labeled with radioactive antibodies, was found to be enriched in dendrites and cell bodies of neurons in the substantia nigra pars compacta and ventral tegmental area. This pattern makes sense for a protein that regulates dopamine levels in the synapse.
Staining in the striatum and nucleus accumbens of the mesolimbic pathway was dense and heterogeneous. In the striatum, DAT is localized in the plasma membrane of axon terminals. Double immunocytochemistry demonstrated DAT colocalization with two other markers of nigrostriatal terminals, tyrosine hydroxylase and D2 dopamine receptors. The latter was thus demonstrated to be an autoreceptor on cells that release dopamine. TAAR1 is a presynaptic intracellular receptor that is also colocalized with DAT and which has the opposite effect of the D2 autoreceptor when activated;[15][25] i.e., it internalizes dopamine transporters and induces efflux through reversed transporter function via PKA and PKC signaling.
Surprisingly, DAT was not identified within any synaptic active zones. These results suggest that striatal dopamine reuptake may occur outside of synaptic specializations once dopamine diffuses from the synaptic cleft.
Within the perikarya of pars compacta neurons, DAT was localized primarily to rough and smooth endoplasmic reticulum, Golgi complex, and multivesicular bodies, identifying probable sites of synthesis, modification, transport, and degradation.[27]
Genetics and regulation
The gene for DAT, known as DAT1, is located on chromosome 5p15.[6] The protein encoding region of the gene is over 64 kb long and comprises 15 coding segments or exons.[28] This gene has a variable number tandem repeat (VNTR) at the 3' end (rs28363170) and another in the intron 8 region.[29] Differences in the VNTR have been shown to affect the basal level of expression of the transporter; consequently, researchers have looked for associations with dopamine-related disorders.[30]
While transcription factors control which cells express DAT, functional regulation of this protein is largely accomplished by kinases. MAPK,[32]CAMKII,[21][22]PKA,[15] and PKC[22][33] can modulate the rate at which the transporter moves dopamine or cause the internalization of DAT. Co-localized TAAR1 is an important regulator of the dopamine transporter that, when activated, phosphorylates DAT through protein kinase A (PKA) and protein kinase C (PKC) signaling.[15][34]Phosphorylation by either protein kinase can result in DAT internalization (non-competitive reuptake inhibition), but PKC-mediated phosphorylation alone induces reverse transporter function (dopamine efflux).[15][35]Dopamine autoreceptors also regulate DAT by directly opposing the effect of TAAR1 activation.[15]
The rate at which DAT removes dopamine from the synapse can have a profound effect on the amount of dopamine in the cell. This is best evidenced by the severe cognitive deficits, motor abnormalities, and hyperactivity of mice with no dopamine transporters.[40] These characteristics have striking similarities to the symptoms of ADHD.
Differences in the functional VNTR have been identified as risk factors for bipolar disorder[41] and ADHD.[42][43] Data has emerged that suggests there is also an association with stronger withdrawal symptoms from alcoholism, although this is a point of controversy.[44][45] An allele of the DAT gene with normal protein levels is associated with non-smoking behavior and ease of quitting.[46] Additionally, male adolescents particularly those in high-risk families (ones marked by a disengaged mother and absence of maternal affection) who carry the 10-allele VNTR repeat show a statistically significant affinity for antisocial peers.[47][48]
Increased activity of DAT is associated with several different disorders, including clinical depression.[49]
Cocaine blocks DAT by binding directly to the transporter and reducing the rate of transport.[12] In contrast, amphetamine enters the presynaptic neuron directly through the neuronal membrane or through DAT, competing for reuptake with dopamine. Once inside, it binds to TAAR1 or enters synaptic vesicles through VMAT2. When amphetamine binds to TAAR1, it reduces the firing rate of the postsynaptic neuron and triggers protein kinase A and protein kinase C signaling, resulting in DAT phosphorylation. Phosphorylated DAT then either operates in reverse or withdraws into the presynaptic neuron and ceases transport. When amphetamine enters the synaptic vesicles through VMAT2, dopamine is released into the cytosol.[15][16] Amphetamine also produces dopamine efflux through a second TAAR1-independent mechanism involving CAMKIIα-mediated phosphorylation of the transporter, which putatively arises from the activation of DAT-coupled L-type calcium channels by amphetamine.[11]
The dopaminergic mechanisms of each drug are believed to underlie the pleasurable feelings elicited by these substances.[7]
Interactions
Dopamine transporter has been shown to interact with:
Apart from these innate protein-protein interactions, recent studies demonstrated that viral proteins such as HIV-1Tat protein interacts with the DAT[58][59] and this binding may alter the dopamine homeostasis in HIV positive individuals which is a contributing factor for the HIV-associated neurocognitive disorders.[60]
↑"Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
↑"Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
↑Carboni E, Tanda GL, Frau R, Di Chiara G (September 1990). "Blockade of the noradrenaline carrier increases extracellular dopamine concentrations in the prefrontal cortex: evidence that dopamine is taken up in vivo by noradrenergic terminals". Journal of Neurochemistry. 55 (3): 1067–70. doi:10.1111/j.1471-4159.1990.tb04599.x. PMID2117046. S2CID23682303.
12Vandenbergh DJ, Persico AM, Hawkins AL, Griffin CA, Li X, Jabs EW, etal. (December 1992). "Human dopamine transporter gene (DAT1) maps to chromosome 5p15.3 and displays a VNTR". Genomics. 14 (4): 1104–6. doi:10.1016/S0888-7543(05)80138-7. hdl:11380/1250902. PMID1478653.
12Wheeler DD, Edwards AM, Chapman BM, Ondo JG (August 1993). "A model of the sodium dependence of dopamine uptake in rat striatal synaptosomes". Neurochemical Research. 18 (8): 927–36. doi:10.1007/BF00998279. PMID8371835. S2CID42196576.
12345Cameron KN, Solis E, Ruchala I, De Felice LJ, Eltit JM (November 2015). "Amphetamine activates calcium channels through dopamine transporter-mediated depolarization". Cell Calcium. 58 (5): 457–66. doi:10.1016/j.ceca.2015.06.013. PMC4631700. PMID26162812. One example of interest is CaMKII, which has been well characterized as an effector of Ca2+ currents downstream of L-type Ca2+ channels [21,22]. Interestingly, DAT is a CaMKII substrate and phosphorylated DAT favors the reverse transport of dopamine [48,49], constituting a possible mechanism by which electrical activity and L-type Ca2+ channels may modulate DAT states and dopamine release.... In summary, our results suggest that pharmacologically, S(+)AMPH is more potent than DA at activating hDAT-mediated depolarizing currents, leading to L-type Ca2+ channel activation, and the S(+)AMPH-induced current is more tightly coupled than DA to open L-type Ca2+ channels.
12Quintero J, Gutiérrez-Casares JR, Álamo C (11 August 2022). "Molecular Characterisation of the Mechanism of Action of Stimulant Drugs Lisdexamfetamine and Methylphenidate on ADHD Neurobiology: A Review". Neurology and Therapy. 11 (4): 1489–1517. doi:10.1007/s40120-022-00392-2. PMC9588136. PMID35951288. The active form of the drug has a central nervous system stimulating activity by the primary inhibition of DAT, NET, trace amine-associated receptor 1 (TAAR1) and vesicular monoamine transporter 2 (SLC18A2), among other targets, therefore regulating the reuptake and release of catecholamines (primarily NE and DA) on the synaptic cleft.... LDX can also promote the increase of DA in the synaptic cleft by activating protein TAAR1, which produces the efflux of monoamine NTs, mainly DA, from storage sites on presynaptic neurons. TAAR1 activation leads to intracellular cAMP signalling that results in PKA and PKC phosphorylation and activation. This PKC activation decreases DAT1, NET1 and SERT cell surface expression, intensifying the direct blockage of monoamine transporters by LDX and improving the neurotransmission imbalance in ADHD.
↑Sulzer D, Cragg SJ, Rice ME (August 2016). "Striatal dopamine neurotransmission: regulation of release and uptake". Basal Ganglia. 6 (3): 123–148. doi:10.1016/j.baga.2016.02.001. PMC4850498. PMID27141430. Despite the challenges in determining synaptic vesicle pH, the proton gradient across the vesicle membrane is of fundamental importance for its function. Exposure of isolated catecholamine vesicles to protonophores collapses the pH gradient and rapidly redistributes transmitter from inside to outside the vesicle.... Amphetamine and its derivatives like methamphetamine are weak base compounds that are the only widely used class of drugs known to elicit transmitter release by a non-exocytic mechanism. As substrates for both DAT and VMAT, amphetamines can be taken up to the cytosol and then sequestered in vesicles, where they act to collapse the vesicular pH gradient.
↑Ledonne A, Berretta N, Davoli A, Rizzo GR, Bernardi G, Mercuri NB (July 2011). "Electrophysiological effects of trace amines on mesencephalic dopaminergic neurons". Front. Syst. Neurosci. 5: 56. doi:10.3389/fnsys.2011.00056. PMC3131148. PMID21772817. Three important new aspects of TAs action have recently emerged: (a) inhibition of firing due to increased release of dopamine; (b) reduction of D2 and GABAB receptor-mediated inhibitory responses (excitatory effects due to disinhibition); and (c) a direct TA1 receptor-mediated activation of GIRK channels which produce cell membrane hyperpolarization.
↑"TAAR1". GenAtlas. University of Paris. 28 January 2012. Retrieved 29 May 2014. • tonically activates inwardly rectifying K(+) channels, which reduces the basal firing frequency of dopamine (DA) neurons of the ventral tegmental area (VTA)
123Vaughan RA, Foster JD (September 2013). "Mechanisms of dopamine transporter regulation in normal and disease states". Trends in Pharmacological Sciences. 34 (9): 489–96. doi:10.1016/j.tips.2013.07.005. PMC3831354. PMID23968642. AMPH and METH also stimulate DA efflux, which is thought to be a crucial element in their addictive properties [80], although the mechanisms do not appear to be identical for each drug [81]. These processes are PKCβ– and CaMK–dependent [72, 82], and PKCβ knock-out mice display decreased AMPH-induced efflux that correlates with reduced AMPH-induced locomotion [72].
↑Maguire JJ, Davenport AP (19 July 2016). "Trace amine receptor: TA1 receptor". IUPHAR/BPS Guide to PHARMACOLOGY. International Union of Basic and Clinical Pharmacology. Retrieved 22 September 2016.
↑Kawarai T, Kawakami H, Yamamura Y, Nakamura S (August 1997). "Structure and organization of the gene encoding human dopamine transporter". Gene. 195 (1): 11–8. doi:10.1016/S0378-1119(97)00131-5. PMID9300814.
↑Sano A, Kondoh K, Kakimoto Y, Kondo I (May 1993). "A 40-nucleotide repeat polymorphism in the human dopamine transporter gene". Human Genetics. 91 (4): 405–6. doi:10.1007/BF00217369. PMID8500798. S2CID39416578.
↑Krause J (April 2008). "SPECT and PET of the dopamine transporter in attention-deficit/hyperactivity disorder". Expert Review of Neurotherapeutics. 8 (4): 611–25. doi:10.1586/14737175.8.4.611. PMID18416663. S2CID24589993. Zinc binds at... extracellular sites of the DAT [103], serving as a DAT inhibitor. In this context, controlled double-blind studies in children are of interest, which showed positive effects of zinc [supplementation] on symptoms of ADHD [105,106]. It should be stated that at this time [supplementation] with zinc is not integrated in any ADHD treatment algorithm.
↑Sulzer D (February 2011). "How addictive drugs disrupt presynaptic dopamine neurotransmission". Neuron. 69 (4): 628–49. doi:10.1016/j.neuron.2011.02.010. PMC3065181. PMID21338876. They did not confirm the predicted straightforward relationship between uptake and release, but rather that some compounds including AMPH were better releasers than substrates for uptake. Zinc, moreover, stimulates efflux of intracellular [3H]DA despite its concomitant inhibition of uptake (Scholze et al., 2002).
12Scholze P, Nørregaard L, Singer EA, Freissmuth M, Gether U, Sitte HH (June 2002). "The role of zinc ions in reverse transport mediated by monoamine transporters". The Journal of Biological Chemistry. 277 (24): 21505–13. doi:10.1074/jbc.M112265200. PMID11940571. The human dopamine transporter (hDAT) contains an endogenous high affinity Zn2+ binding site with three coordinating residues on its extracellular face (His193, His375, and Glu396).... Although Zn2+ inhibited uptake, Zn2+ facilitated [3H]MPP+ release induced by amphetamine, MPP+, or K+-induced depolarization specifically at hDAT but not at the human serotonin and the norepinephrine transporter (hNET).
↑Scassellati C, Bonvicini C, Faraone SV, Gennarelli M (October 2012). "Biomarkers and attention-deficit/hyperactivity disorder: a systematic review and meta-analyses". Journal of the American Academy of Child and Adolescent Psychiatry. 51 (10): 1003–1019.e20. doi:10.1016/j.jaac.2012.08.015. PMID23021477. With regard to zinc supplementation, a placebo controlled trial reported that doses up to 30mg/day of zinc were safe for at least 8weeks, but the clinical effect was equivocal except for the finding of a 37% reduction in amphetamine optimal dose with 30mg per day of zinc.110
↑Greenwood TA, Alexander M, Keck PE, McElroy S, Sadovnick AD, Remick RA, etal. (March 2001). "Evidence for linkage disequilibrium between the dopamine transporter and bipolar disorder". American Journal of Medical Genetics. 105 (2): 145–51. doi:10.1002/1096-8628(2001)9999:9999<::AID-AJMG1161>3.0.CO;2-8. PMID11304827.
↑Yang B, Chan RC, Jing J, Li T, Sham P, Chen RY (June 2007). "A meta-analysis of association studies between the 10-repeat allele of a VNTR polymorphism in the 3'-UTR of dopamine transporter gene and attention deficit hyperactivity disorder". American Journal of Medical Genetics. Part B, Neuropsychiatric Genetics. 144B (4): 541–50. doi:10.1002/ajmg.b.30453. PMID17440978. S2CID22881996.
↑Sander T, Harms H, Podschus J, Finckh U, Nickel B, Rolfs A, etal. (February 1997). "Allelic association of a dopamine transporter gene polymorphism in alcohol dependence with withdrawal seizures or delirium". Biological Psychiatry. 41 (3): 299–304. doi:10.1016/S0006-3223(96)00044-3. PMID9024952. S2CID42947314.
↑Ueno S (February 2003). "Genetic polymorphisms of serotonin and dopamine transporters in mental disorders". The Journal of Medical Investigation. 50 (1–2): 25–31. PMID12630565.
↑Beaver KM, Wright JP, DeLisi M (September 2008). "Delinquent peer group formation: evidence of a gene x environment correlation". The Journal of Genetic Psychology. 169 (3): 227–44. doi:10.3200/GNTP.169.3.227-244. PMID18788325. S2CID46592146.
↑Kostrzewa RM (2022). "Survey of Selective Monoaminergic Neurotoxins Targeting Dopaminergic, Noradrenergic, and Serotoninergic Neurons". Handbook of Neurotoxicity. Cham: Springer International Publishing. pp.159–198. doi:10.1007/978-3-031-15080-7_53. ISBN978-3-031-15079-1.
↑Storch A, Ludolph AC, Schwarz J (October 2004). "Dopamine transporter: involvement in selective dopaminergic neurotoxicity and degeneration". J Neural Transm (Vienna). 111 (10–11): 1267–1286. doi:10.1007/s00702-004-0203-2. PMID15480838.
↑Sotnikova TD, Beaulieu JM, Gainetdinov RR, Caron MG (February 2006). "Molecular biology, pharmacology and functional role of the plasma membrane dopamine transporter". CNS Neurol Disord Drug Targets. 5 (1): 45–56. doi:10.2174/187152706784111579. PMID16613553.
↑Aceto MD, Botton I, Martin R, Levitt M, Bentley HC, Speight PT (1970). "Pharmacologic properties and mechanism of action of amfonelic acid". Eur J Pharmacol. 10 (3): 344–354. doi:10.1016/0014-2999(70)90206-2. PMID4393073.
↑Porrino LJ, Goodman NL, Sharpe LG (November 1988). "Intravenous self-administration of the indirect dopaminergic agonist amfonelic acid by rats". Pharmacol Biochem Behav. 31 (3): 623–626. doi:10.1016/0091-3057(88)90240-7. PMID2908003.
↑Garattini S (July 1997). "Pharmacology of amineptine, an antidepressant agent acting on the dopaminergic system: a review". Int Clin Psychopharmacol. 12 (Suppl 3): S15 –S19. doi:10.1097/00004850-199707003-00003. PMID9347388.
↑Bonnet JJ, Chagraoui A, Protais P, Costentin J (1987). "Interactions of amineptine with the neuronal dopamine uptake system: neurochemical in vitro and in vivo studies". J Neural Transm. 69 (3–4): 211–220. doi:10.1007/BF01244342. PMID3625193.
↑Dutta AK, Zhang S, Kolhatkar R, Reith ME (October 2003). "Dopamine transporter as target for drug development of cocaine dependence medications". Eur J Pharmacol. 479 (1–3): 93–106. doi:10.1016/j.ejphar.2003.08.060. PMID14612141.
↑Katz JL, Newman AH, Izenwasser S (July 1997). "Relations between heterogeneity of dopamine transporter binding and function and the behavioral pharmacology of cocaine". Pharmacol Biochem Behav. 57 (3): 505–512. doi:10.1016/s0091-3057(96)00441-8. PMID9218275.
↑Bergman J, France CP, Holtzman SG, Katz JL, Koek W, Stephens DN (December 2000). "Agonist efficacy, drug dependence, and medications development: preclinical evaluation of opioid, dopaminergic, and GABAA-ergic ligands". Psychopharmacology (Berl). 153 (1): 67–84. doi:10.1007/s002130000567. PMID11255930.
↑Izenwasser S, Newman AH, Cox BM, Katz JL (February 1996). "The cocaine-like behavioral effects of meperidine are mediated by activity at the dopamine transporter". Eur J Pharmacol. 297 (1–2): 9–17. doi:10.1016/0014-2999(95)00696-6. PMID8851160.
123Golovko AI, Bonitenko EY, Ivanov MB, Barinov VA, Zatsepin EP (2016). "The neurochemical bases of the pharmacological activity of ligands of monoamine-transport systems". Neurochemical Journal. 10 (3): 173–183. doi:10.1134/S1819712416030065. ISSN1819-7124.
↑Hauck Newman A, Katz JL (2008). "Atypical Dopamine Uptake Inhibitors that Provide Clues About Cocaine's Mechanism at the Dopamine Transporter". Transporters as Targets for Drugs. Topics in Medicinal Chemistry. Vol.4. Berlin, Heidelberg: Springer Berlin Heidelberg. pp.95–129. doi:10.1007/7355_2008_027. ISBN978-3-540-87911-4.
↑Naglich AC, Brown ES, Adinoff B (2019). "Systematic review of preclinical, clinical, and post-marketing evidence of bupropion misuse potential". Am J Drug Alcohol Abuse. 45 (4): 341–354. doi:10.1080/00952990.2018.1545023. PMID30601027.
↑Noe G, Shah K, Ongchuan S, Munjal S (2024). "Clinical Presentations of Bupropion Prescription Drug Misuse: A Systematic Review". J Clin Psychopharmacol. 44 (3): 284–290. doi:10.1097/JCP.0000000000001858. PMID38656298.
123Rothman RB, Glowa JR (1995). "A review of the effects of dopaminergic agents on humans, animals, and drug-seeking behavior, and its implications for medication development. Focus on GBR 12909". Mol Neurobiol. 11 (1–3): 1–19. doi:10.1007/BF02740680. PMID8561954.
123Salamone JD, Correa M (January 2024). "The Neurobiology of Activational Aspects of Motivation: Exertion of Effort, Effort-Based Decision Making, and the Role of Dopamine". Annu Rev Psychol. 75: 1–32. doi:10.1146/annurev-psych-020223-012208. hdl:10234/207207. PMID37788571.
↑Böning J, Fuchs G (September 1986). "Nomifensine and psychological dependence--a case report". Pharmacopsychiatry. 19 (5): 386–388. doi:10.1055/s-2007-1017275. PMID3774872.
↑Veinberg G, Vavers E, Orlova N, Kuznecovs J, Domracheva I, Vorona M, etal. (2015). "Stereochemistry of phenylpiracetam and its methyl derivative: improvement of the pharmacological profile". Chemistry of Heterocyclic Compounds. 51 (7): 601–606. doi:10.1007/s10593-015-1747-9. ISSN0009-3122. Phenylpiracetam was originally designed as a nootropic drug for the sustenance and improvement of the physical condition and cognition abilities of Soviet space crews.2 Later, especially during the last decade, phenylpiracetam was introduced into general clinical practice in Russia and in some Eastern European countries. The possible target receptors and mechanisms for the acute activity of this drug remained unclear, until very recently it was found that (R)-phenylpiracetam (5) (MRZ-9547) is a selective dopamine transporter inhibitor that moderately stimulates striatal dopamine release.19
12Sommer S, Danysz W, Russ H, Valastro B, Flik G, Hauber W (December 2014). "The dopamine reuptake inhibitor MRZ-9547 increases progressive ratio responding in rats". The International Journal of Neuropsychopharmacology. 17 (12): 2045–2056. doi:10.1017/S1461145714000996. PMID24964269. Here, we tested the effects of MRZ-9547 [...], and its l-enantiomer MRZ-9546 on effort-related decision making in rats. The racemic form of these compounds referred to as phenotropil has been shown to stimulate motor activity in rats (Zvejniece et al., 2011) and enhance physical capacity and cognition in humans (Malykh and Sadaie, 2010). [...] MRZ-9547 turned out to be a DAT inhibitor as shown by displacement of binding of [125I] RTI-55 (IC50 = 4.82 ± 0.05 μM, n=3) to human recombinant DAT expressed in CHO-K1 cells and inhibition of DA uptake (IC50 = 14.5 ± 1.6 μM, n=2) in functional assays in the same cells. It inhibited norepinephrine transporter (NET) with an IC50 of 182 μM (one experiment in duplicate). The potencies for the l-enantiomer MRZ-9546 were as follows: DAT binding (Ki = 34.8 ± 14.8 μM, n=3), DAT function (IC50 = 65.5 ± 8.3 μM, n=2) and NET function (IC50 = 667 μM, one experiment performed in duplicate).
↑Zvejniece L, Svalbe B, Vavers E, Makrecka-Kuka M, Makarova E, Liepins V, etal. (September 2017). "S-phenylpiracetam, a selective DAT inhibitor, reduces body weight gain without influencing locomotor activity". Pharmacology, Biochemistry, and Behavior. 160: 21–29. doi:10.1016/j.pbb.2017.07.009. PMID28743458. S2CID13658335.
↑Stutz PV, Golani LK, Witkin JM (February 2019). "Animal models of fatigue in major depressive disorder". Physiology & Behavior. 199: 300–305. doi:10.1016/j.physbeh.2018.11.042. PMID30513290. In a study performed by Sommer et al. (2014), healthy rats treated with the selective dopamine transport (DAT) inhibitor MRZ-9547 (Fig. 1) chose high effort, high reward more often than their untreated matched controls. Unlike similar studies, however, depressive symptoms were not induced before treatment; rather, baseline healthy controls were compared to healthy rats treated with MRZ-9547. [...] In one study, the selective DAT inhibitor MRZ-9547 increased the number of lever presses more than untreated controls (Sommer et al., 2014). The investigators concluded that such effort-based "decision making in rodents could provide an animal model for motivational dysfunctions related to effort expenditure such as fatigue, e.g. in Parkinson's disease or major depression." Based upon the findings with MRZ-9547, they suggested that this drug mechanism might be a valuable therapeutic entity for fatigue in neurological and neuropsychiatric disorders. [...] A high effort bias been reported with bupropion (Randall et al., 2015), lisdexamfetamine (Yohn etal., 2016e), and the DA uptake blockers MRZ-9547 (Sommer et al., 2014), PRX-14040 (Fig. 1) (Yohn et al., 2016d) and GBR12909 (Fig. 1) (Yohn et al., 2016c).
↑Dekundy A, Mela F, Hofmann M, Danysz W (June 2015). "Effects of dopamine uptake inhibitor MRZ-9547 in animal models of Parkinson's disease". Journal of Neural Transmission. 122 (6): 809–818. doi:10.1007/s00702-014-1326-8. PMID25319446.
↑Ngo Q, Plante DT (19 September 2022). "An Update on the Misuse and Abuse Potential of Pharmacological Treatments for Central Disorders of Hypersomnolence". Current Sleep Medicine Reports. 8 (4): 147–159. doi:10.1007/s40675-022-00227-4. ISSN2198-6401.
↑Yang J, Gao J (August 2019). "Solriamfetol for the treatment of excessive daytime sleepiness associated with narcolepsy". Expert Rev Clin Pharmacol. 12 (8): 723–728. doi:10.1080/17512433.2019.1632705. PMID31215815.
↑Yohn SE, Gogoj A, Haque A, Lopez-Cruz L, Haley A, Huxley P, etal. (September 2016). "Evaluation of the effort-related motivational effects of the novel dopamine uptake inhibitor PRX-14040". Pharmacol Biochem Behav. 148: 84–91. doi:10.1016/j.pbb.2016.06.004. PMID27296079.
123Rothman RB, Baumann MH (December 2005). "Targeted screening for biogenic amine transporters: potential applications for natural products". Life Sci. 78 (5): 512–518. doi:10.1016/j.lfs.2005.09.001. PMID16202429.
123Rothman RB, Vu N, Partilla JS, Roth BL, Hufeisen SJ, Compton-Toth BA, etal. (October 2003). "In vitro characterization of ephedrine-related stereoisomers at biogenic amine transporters and the receptorome reveals selective actions as norepinephrine transporter substrates". J Pharmacol Exp Ther. 307 (1): 138–145. doi:10.1124/jpet.103.053975. PMID12954796.
↑Hutson PH, Pennick M, Secker R (December 2014). "Preclinical pharmacokinetics, pharmacology and toxicology of lisdexamfetamine: a novel d-amphetamine pro-drug". Neuropharmacology. 87: 41–50. doi:10.1016/j.neuropharm.2014.02.014. PMID24594478.
↑Heal DJ, Cheetham SC, Smith SL (December 2009). "The neuropharmacology of ADHD drugs in vivo: insights on efficacy and safety". Neuropharmacology. 57 (7–8): 608–618. doi:10.1016/j.neuropharm.2009.08.020. PMID19761781.
↑Fuller RW, Perry KW, Bymaster FP, Wong DT (March 1978). "Comparative effects of pemoline, amfonelic acid and amphetamine on dopamine uptake and release in vitro and on brain 3,4-dihydroxyphenylacetic acid concentration in spiperone-treated rats". The Journal of Pharmacy and Pharmacology. 30 (3): 197–198. doi:10.1111/j.2042-7158.1978.tb13201.x. PMID24701.
12Zhao G, Qin GW, Wang J, Chu WJ, Guo LH (January 2010). "Functional activation of monoamine transporters by luteolin and apigenin isolated from the fruit of Perilla frutescens (L.) Britt". Neurochem Int. 56 (1): 168–176. doi:10.1016/j.neuint.2009.09.015. PMID19815045.
↑Zhang J, Liu X, Lei X, Wang L, Guo L, Zhao G, etal. (November 2010). "Discovery and synthesis of novel luteolin derivatives as DAT agonists". Bioorg Med Chem. 18 (22): 7842–7848. doi:10.1016/j.bmc.2010.09.049. PMID20971650.
↑Rothman RB, Dersch CM, Carroll FI, Ananthan S (March 2002). "Studies of the biogenic amine transporters. VIII: identification of a novel partial inhibitor of dopamine uptake and dopamine transporter binding". Synapse. 43 (4): 268–274. doi:10.1002/syn.10046. PMID11835522.
↑Al'tshuler RA (2005). "Comparative Molecular Model Estimation of the Affinity of Phenylethylamines to the Binding Sites of Membrane Transporters". Pharmaceutical Chemistry Journal. 39 (4): 169–175. doi:10.1007/s11094-005-0110-3. ISSN0091-150X.
↑Harsing LG, Sershen H, Lajtha A (1994). "Evidence that ibogaine releases dopamine from the cytoplasmic pool in isolated mouse striatum". J Neural Transm Gen Sect. 96 (3): 215–225. doi:10.1007/BF01294788. PMID7826572.
Overview of all the structural information available in the PDB for UniProt: Q7K4Y6 (Drosophila melanogaster Sodium-dependent dopamine transporter) at the PDBe-KB.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.